Introduction
Peroxiredoxins (Prxs) have recently been the subject of growing interest as a new family of thiol-specific antioxidant proteins [1]. These agents have been shown to play a critical role in peroxide detoxification by using reducing equivalents provided through the thioredoxin system but not from glutaredoxin [2,3]. They exist in multiple isoforms [4,5] and catalyze the reduction of a broad range of different peroxides, including H2O2, alkyl hydroperoxides and peroxynitrite. The existence of different members of the Prx family has already been observed in a variety of organisms, ranging from archaea to mammals [6].
The mammalian peroxiredoxin family consists of six proteins (Prx I-VI) expressed as unique gene products, with the capability of reducing hydrogen peroxide, lipid hydroperoxides, and peroxynitrite [7-10]. Prx I-IV are categorized as typical 2-Cys peroxiredoxins, distinct from the atypical 2-Cys (Prx V) and 1-Cys peroxiredoxin (Prx VI). Prx I-IV are members of a family containing a cysteine (Cys) on both the N- and C-terminal and form a homodimer with a head-to-tail type association [2]. Prx V, a monomeric form, also contains two Cys; this protein is the most recently discovered Prx from a mammalian system [11]. Prx V is known to regulate apoptosis by inhibiting the p53-induced reactive oxygen species (ROS) formation [12]. Prx VI (1-Cys) contains only one Cys residue in the catalytic site and functions as a monomer [2]. Among these Prxs, Prx I is a basic (pH 8.3) 22-kDa protein localized to the cytosol. Prx I is upregulated in a variety of cell types following exposure to oxidative stress. Upregulation of Prx I in response to H2O2 was first demonstrated in mouse peritoneal macrophages [13] and in cultured vascular smooth muscle cells following exposure to oxidized low-density lipoprotein [14].
In the ovary, regression of the corpus luteum is an example of degeneration of cellular function. Luteinizing hormone (LH) causes depletion of ascorbic acid in the corpus luteum. Furthermore, reactive oxygen species have been shown to play a role in the regression of the corpus luteum [15]. However, the expression of Prx I in the ovary has not been reported. In the present study, therefore, the expression of Prx I was examined in the rat ovary.
Methods
1. Animals
Immature female rats of the Sprague-Dawley strain were purchased from Daehan Laboratories (Eumsung, Korea). They were housed in groups in a room with a controlled temperature and photoperiod (10-hours dark, 14-hours light, with lights on from 0600-2000 hours). The animals had access to food and water ad libitum. The animals, ranging in age from 3-21 days, were killed by cervical dislocation, and the ovaries were removed for RNA analysis. Ovaries were also collected from immature (26-day-old) rats at various times after treatment with 10 IU pregnant mare's serum gonadotropin (PMSG) (Sigma Chemical Co., St. Louis, MO, USA) to induce multiple follicle growth. To obtain immature growing granulosa cells for the proliferation assay, rats were treated with diethylstilbestrol (DES) (Sigma Chemical Co.) at 22 days of age. Some rats received a single intraperitoneal injection of 10 IU human chorionic gonadotropin (hCG) (Sigma Chemical Co.) to induce ovulation, and ovaries were obtained at different time intervals for northern blot and in situ hybridization analysis. Three to four ovarian samples obtained from different animals were included in the analysis. All animal handling and procedures were approved by the Institutional Animal Care and Use Committee of Chonnam National University.
2. PCR cloning of rat Prx I
Total RNA from the rat brain, testis, and lung was isolated using Tri-reagent solution. Five micrograms of total RNA were reverse transcribed using the Superscript Preamplification System (Gibco, Gaithersburg, MD, USA) according to the instruction manual. One microgram of reverse transcribed cDNA and 100 pM of primer (Prx I's antisense, 5'-GAG TTT CTT AAA TTC TTC TGC TCT A-3' and sense, 5'-CTT CAG GAA ATG CAA AAA TTG GGC AT-3') was added in a final volume of 100 µL containing 250 µM each deoxynucleotide triphosphates, 2 mM MgCl2, and 0.2 U Taq polymerase. PCR was performed for 30 cycles with denaturing at 94℃ for 1 minute, annealing at 65℃ for 1.5 minutes, and elongation at 72℃ for 1 minute. PCR products of the expected size (Prx I; 203 bp) were subcloned into pGEM-T Easy Vector (Promega Co., Madison, WI, USA), and sequenced using T7 and T3 primers with automatic DNA sequencer (Perkin-Elmer, Foster City, CA, USA).
3. Northern blot analysis
Total RNA from ovaries was isolated using Tri-reagent solution (Molecular Research Center Inc., Cincinnati, OH, USA). Twenty micrograms of total RNA were fractionated by electrophoresis on a 1% agarose gel containing formaldehyde, and transferred to nylon membranes by capillary blotting with 10×sodium citrate-sodium chloride (SSC). After UV cross-linking and prehybridization, membranes were hybridized overnight at 42℃ in solution containing 50% formaldehyde, 5×SSC, 1 mM EDTA, 250 µg/mL denatured salmon sperm DNA, and a total of 2-4×106 cpm of α32P-labeled rat Prx I cDNA probes. After hybridization, membranes were washed twice for 5 minutes at room temperature in 2×SSC and 0.1% SDS, followed by 1 hour at 65℃ in 0.5×SSC and 0.1% SDS. Membranes were then exposed using Kodak RX films (Eastman Kodak Co., Rochester, NY, USA) for 1-4 days at -80℃.
4. In situ hybridization analysis
Rat ovaries were fixed at 4℃ for 6 hours in 4% paraformaldehyde in phosphate-buffered saline (PBS), followed by immersion in 0.5 M sucrose in PBS overnight. Cryostat sections (14 µm thick) were mounted on poly-L-lysine (Sigma Chemical Co.) coated microscope slides, fixed in 4% paraformaldehyde in PBS, and stored at -70℃ until analyzed. The hybridization procedure was essentially the same as previously described [16]. In brief, sections were pretreated serially with 0.2 M HCl, 2×SSC, pronase E (0.125 mg/mL), 4% paraformaldehyde, and acetic anhydride in triethanolamine. Hybridization was carried out at 52-55℃ overnight in the mixture containing 35S-labeled rat Prx I cRNA probe (108 cpm/mL), 50% formamide, 0.3 M NaCl, 10 mM Tris-HCl, 5 mM EDTA, 1×Denhardt's solution, 10% dextran sulfate, 1 mg/mL carrier transfer RNA, and 10 mM dithiothreitol. Post-hybridization washing was performed under stringent conditions that included ribonuclease A (25 mg/mL) treatment at 37℃ for 30 minutes and a final stringency of 0.1×SSC. Slides were dipped into NTB-2 emulsion (Eastman Kodak Co.) and exposed at 4℃ until being developed after 3 weeks. The slides were stained with hematoxylin and eosin and examined under a light microscope with bright- and dark-field illumination.
Results
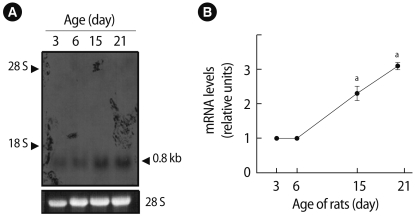
1. Levels of Prx I mRNAs during development
The developmental changes in Prx I mRNA levels in the ovary were determined by a northern blot analysis. As shown in Figure 1A, the levels of 0.8 Kb Prx I transcripts were increased during development. The levels of ovarian Prx I transcript were three times higher in 21-day-old rats and two times higher in 15-day-old rats than those in 3-day-old or 6-day-old rats (p<0.05) (Figure 1B).
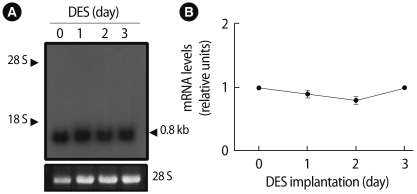
2. Regulation of Prx I expression by estrogen
To study the effect of estrogen on Prx I expression in vivo, total RNA extracted from ovaries at different days after implantation of capsules containing diethylstilbestrol (DES) was analyzed by northern blotting. The levels of Prx I mRNA slightly, but not significantly, declined 1-2 days after DES treatment (Figure 2).
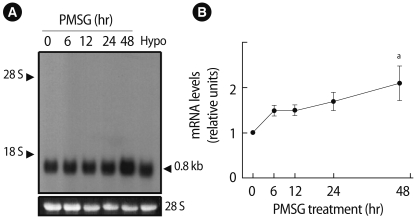
3. Gonadotropin regulation of Prx I mRNA levels
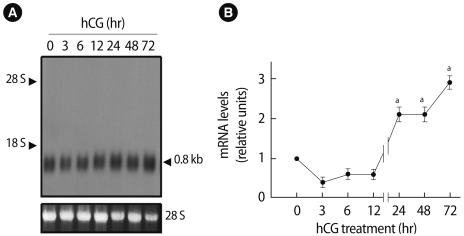
To examine gonadotropin regulation of Prx I mRNA levels in the ovary, total RNA extracted from ovaries at different times after gonadotropin treatments was analyzed by northern blotting. As shown in Figure 3A, PMSG treatment of 26-day-old immature rats caused an increase in ovarian expression of the Prx I gene. Prx I expression was markedly increased in ovaries treated with PMSG for 48 hours (two-fold increase; p<0.05) (Figure 3B). The hypothalamus used as a positive control also expressed the Prx I gene. Furthermore, treatment with hCG of PMSG-primed rats resulted in the stimulation of Prx I mRNAs within 24 hours (p<0.05) (Figure 4A). High levels remained until 72 hours post-hCG treatment, which corresponds to the period of corpus luteum formation (Figure 4B). The levels of Prx I mRNA slightly declined 9-12 hours after stimulation.
4. Localization of Prx I mRNAs in the rat ovary
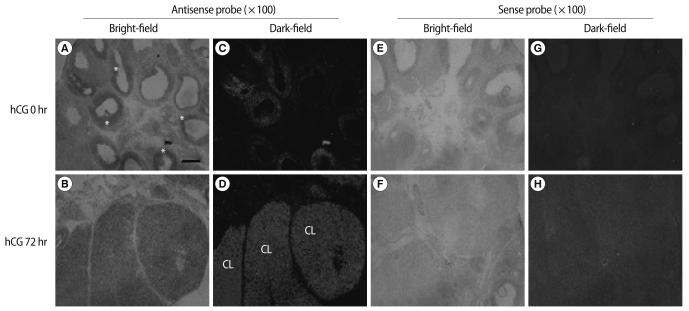
To determine the cell types expressing Prx I mRNA in hormone-stimulated ovaries, antisense and sense cRNA probes for rat Prx I were generated for in situ hybridization analysis. In the ovaries of PMSG-primed immature rats, Prx I transcripts were detected in granulosa cells of preovulatory follicles (Figure 5A, 5C). In addition, in the ova-ries of PMSG-primed immature rats followed by hCG stimulation for 72 hours, high levels of Prx I mRNA were expressed in corpus luteum cells (Figure 5B, 5D). No specific signal was detected in follicles hybridized with the sense probe (Figure 5E-5H).
Discussion
The present study demonstrates that Prx I levels were regulated during follicle development in the rat ovary. Northern blot analysis, showing an increase in 0.8 kb Prx I transcription in the ovaries of 21-day-old rat, suggests the possible role of Prx in follicle growth. Furthermore, the expression of Prx isotypes was stimulated by gonadotropins during corpus luteum formation.
Reactive oxygen species (ROS), the natural metabolites of intracellular oxygen, are constantly created and destroyed within the cell as a means of maintaining strict redox homeostasis. ROS such as hydrogen peroxide (H2O2) and superoxide are created as a result of normal cellular signaling and metabolism [17], including oxidative phosphorylation [18]. Other causes of increased ROS levels include exposure to various drugs and hormones [19] and the overexpression of certain oncoproteins [20]. A group of enzymes that plays a major role in ROS regulation is the Prx family. Prxs reduce hydrogen peroxide and other peroxide substrates via conserved cysteine residues utilizing thiol-containing proteins, such as glutathione or thioredoxin, as electron donors [2,21,22].
Intracellular ROS are a double-edged sword with both beneficial and deleterious consequences. On the one hand, ROS participate in normal cell signaling pathways [22-24]. For example, H2O2 oxidizes cysteine residues in protein-tyrosine phosphatases, which inhibits their function and leads to the upregulation of multiple tyrosine kinase-dependent signaling pathways in response to growth factors [8,25]. The best studied means for regulating ROS levels is via the Prxs. Each of the six mammalian Prxs has a specific tissue and subcellular distribution [2,21]. The most abundant and ubiquitous member of the Prxs, Prx I, is expressed at high levels in virtually every tissue, primarily as a nuclear ROS scavenger. Reactive oxygen species, including the superoxide anion and hydrogen peroxide, are generated in the ovary, but their origin is unknown [26]. Follicles and corpora lutea are subject to ischemic episodes, and leukocytic infiltration is evident at ovulation and leuteolysis [27]. Furthermore, xanthine oxidase in the presence of hypoxanthine evokes marked antigonadotropic and antisteroidogenic actions in ovarian cells [28]. Recent evidence links superoxide radical production to the regulation of corpus luteum function in the rat. In vivo treatment with prostaglandin F2α stimulates a burst of superoxide radicals shortly before progesterone secretion decreases [26], and generation of these radicals appears to be controlled by intracellular signaling pathways [29]. Therefore, the present observation, showing the stimulation of Prx I expression during corpus luteum formation, may implicate Prx in the initiation of corpus luteum formation by modulating superoxide radical production, and in turn, the generated superoxide radicals may alter progesterone production and thereby maintain the formation of the corpus luteum.
During follicle development, Prx I may play an important role in theca cell differentiation. It is known that ovarian follicular growth begins and proceeds to the late preantral stage independently of gonadotropin regulation [30]. Further development depends upon FSH acting upon its cognate receptor expressed by granulosa cells [31]. It has been suggested that insulin-like growth factor I, expressed in granulosa cells of healthy growing follicles, may promote follicle growth by augmenting granulosa cell FSH receptor responsiveness [32], and thereby amplifying FSH-induced aromatase expression and LH receptor induction [33]. Similarly, activin has been shown to promote folliculogenesis during the preantral or early antral stages of growth of follicles by amplifying granulosa cell responsiveness to FSH [34]. Because the pattern of Prx expression seems to follow the spatial expression of P450scc [35] and steroidogenic acute regulatory protein [36] in cells engaged in steroidogenic activity, Prx I expression in the theca cells may be correlated with terminal commitment of cells for steroidogenic differentiation.
In summary, the present study demonstrates the presence and regulation of Prx I expression in the gonadotropin-treated ovaries. Stimulation of Prx I expression by gonadotropins indicates that Prx I may have a physiological function during ovarian follicle development. Particularly, the function of Prx I in corpus luteum formation remains to be established.