Introduction
Recent advances in in vitro culture techniques for human embryos have increased the rate of blastocyst formation. Therefore, in human assisted reproduction, blastocyst transfer is now preferred to the collection of a 3-day-old cleavage stage embryo using the routine method [1,2,3]. This is because it is possible to select a growth-arrested embryo at the 8-cell-stage, and because the transfer of 1 to 2 embryos with a healthy trophectoderm (TE) and inner cell mass (ICM), increases the rates of pregnancy and implantation and reduces the incidence of multiple pregnancies [4,5,6]. The possibility of cryopreserving residual embryos 5 days after embryo transfer increases the likelihood of pregnancy while reducing the time, monetary, and physical burdens of patients struggling with infertility. Multiple studies have explored ways of increasing the rates of viability and hatching of cryopreserved and thawed blastocysts [6,7,8,9].
The cryopreservation of blastocysts using vitrification has been associated with a lower survival rate than observed for fresh embryos, due to the intracellular and extracellular formation of ice crystals caused by incomplete dehydration in the watery environment of the blastocoel cavity and by mixing with vitrifying solution during dehydration [9,10,11]. Moreover, the hatching rate has been found to be decreased by zona hardening, which results from in vitro culture and thawing [9,12,13,14]. The above factors reduce the pregnancy rate in human in vitro fertilization/embryo transfer [15,16,17]. The introduction of artificial shrinkage (AS) and vitrification to prevent the formation of ice crystals inside and outside of the blastocyst increased the survival rate of embryos, but zona hardening during the freezing-thawing processes lowered the hatching rate. Various approaches have been developed to overcome this difficulty [9,17,18,19]. Partial zona dissection (PZD) using a micropipette handled with a micromanipulator has been used, and is promising in that it allows a wide choice of the dissection range. However, since no perivitelline space is present between the TE and the zona pellucida (ZP), this technique pierce TE cells during the assisted hatching (AH) procedure, which may cause some cells to die. Additionally, the outcomes may vary among operators, and the range of dissection in the ZP has not been standardized. Recently, Strohmer and Feichtinger [20] proposed laser-AH (LAH), in which a hole is made in the ZP with a laser beam in order to complete the AH. In this method, the intensity of the laser beam can be controlled, and it is possible to eliminate some wide portions by successively emitting the laser beam onto the ZP. The present study was conducted to characterize the developmental ability of expanded mouse blastocysts depending on whether AH was performed before or after vitrification and thawing (V-T). Another goal of this study was to characterize the effect of AS and AH on the cell number in mouse blastocysts, as assessed by differentially staining TE and ICM with a fluorescent stain after 48 hours of cultivation.
Methods
1. Embryo preparation
1) Animals
ICR mice aged 4 to 5 weeks (female) and 8 to 15 weeks (male) were purchased from the Damool experimental animal center (Daejeon, Korea). All animals were housed with a 12-hour light/dark photoperiod and maintained in an air-conditioned (22℃-25℃) and humidity-controlled (40%-60%) room.
2) Recovery of 2-cell embryos and culture of expanded blastocysts
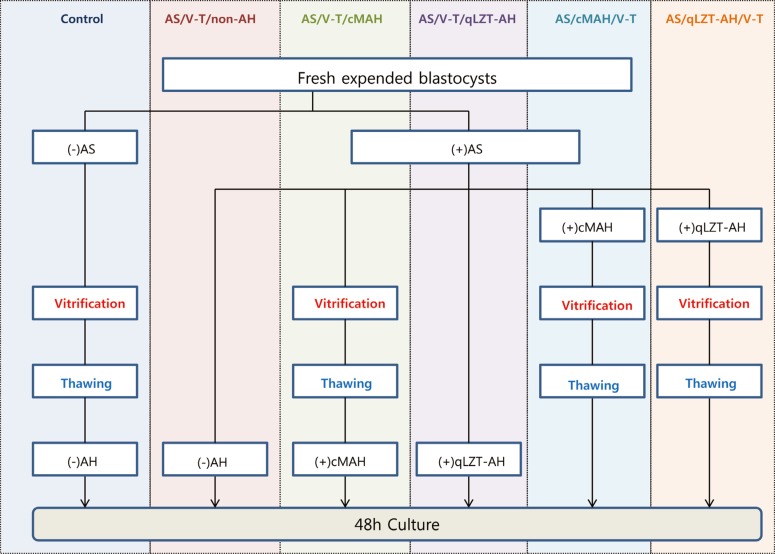
In order to induce superovulation, 7.5 IU of pregnant mare's serum (Sigma, St. Louis, MO, USA) and 7.5 IU of human chorionic gonadotropin (hCG, Sigma) were intraperitoneally injected at an interval of 48 hours. After the injections, female mice were caged with male mice at a 1 to 1 ratio. Females were observed for the presence of a vaginal plug 12 hours later. Females with a vaginal plug were selected and used in this experiment. Animals were sacrificed by cervical dislocation 44 to 48 hours after hCG injection. The oviducts were surgically removed under a dissecting microscope. 2-cell embryos were collected by flushing with the oviducts with 1-mL insulin syringe filled with Ham's F-10 (11-090-1K, Biological lndustries, NY, USA) supplemented with 10% serum substitute supplement (Irvine Scientific, Santa Ana, CA, USA), and cultured in a 60-mm culture dish (3002, Falcon, Santa Ana, CA, USA) with 50-µL of cleavage medium (COOK, Queensland, Australia). A drop of mineral oil (M-3516, Sigma) was used to cover the culture. The embryos were cultured in an incubator for 48 hours at 37℃ and 6% CO2. Expanded blastocysts with a clearly distinguishable ICM and TE were recovered, and randomly allocated into the experimental groups. The expanded blastocysts were placed into two groups. In one group, AS was not conducted (control; non-treatment), and the others were treated with AS. The AS group was further divided into the following subgroups, based on various combinations of V-T, AH, cross-mechanical AH (cMAH), and quarter-laser zona thinning-AH (qLZT-AH): AS/V-T/non-AH (group A, not treated with AS after thawing), AS/V-T/cMAH (group B, treated with cMAH), AS/V-T/qLZT-AH (group C, treated with qLZT-AH), AS/cMAH/V-T (group D, cMAH was performed before freezing and thawing), and AS/qLZT-AH/V-T (group E, treated with AS and qLZT-AH before freezing and thawing) (Figure 1).
2. Artificial shrinkage
After a drop containing 50-µL of blastocyst medium (BM, COOK) was placed in the center of the culture dish (60 mm in diameter) and covered with mineral oil, it was preincubated for more than 6 hours at 37℃ and 6% CO2. Using a micromanipulator (TE2000-U, Nikon, Tokyo, Japan) equipped with an inverted phase-contrast microscope, holding pipette (Sunlight Medical, Jacksonvile, FL, USA), and partial zona dissection pipettes (30°; Sunlight Medical), with tips oriented parallel to each other. AS was carried out in order to artificially dehydrate the blastocoel by piercing through the TE in the ICM, which was oriented to face 12 o'clock.
3. Assisted hatching
1) Cross-mechanical AH
The preparation of the medium and pipettes was the same as in AS. The dissected portion by dissected according to the AS protocol was placed in the 6 o'clock position, fixed with a holding pipette, turned 90°, and dissected in the shape of 'a plus sign', which indicated cruciate thinning of the zona.
2) Quarter-laser zona thinning-AH
LAH was performed under an inverted phase-contrast microscope (×40), which was attached to an RI Saturn 5 Active Laser and Display (B19940, Samsung, Suwon, Korea). The LAH system had a resolution of 1,600×1,200, and it was possible to make a hole with a short wavelength of 1480 nm/400 mW, corresponding to the full laser power routinely used in the image plane and reducing the deformation caused by heat. qLZT-AH was performed after 5 to 10 blastocysts were placed in a drop 50-µL of BM in a 60-mm incubation dish. While observing the blastocysts on the monitor, a virtual line was drawn on the outer rim of the ZP and shaped according to the curved of the ZP. A laser beam was sequentially emitted along this line to make the entire ZP 25% thinner. The average number of emissions was 20 to 30 and the size of the laser holes was 3 to 5 µm. The perforations were located between the center of the ZP and the outer line. Approximately 70% was removed.
4. Vitrification and thawing
All V-T procedures were conducted at room temperature (24℃-26℃). The plain medium used for blastocysts was Dulbecco's phosphate-buffered saline supplemented with 10% serum substitute supplement. The blastocysts were transferred into vitrification solution 1, which was mixed with plain medium containing 7.5% ethylene glycol and 7.5% dimethyl sulfoxide (DMSO) for 10 minutes and then transferred into vitrification solution 2 and mixed with plain medium containing 15% ethylene glycol, 15% DMSO, and 0.5 M sucrose 1 minute. Embryos less than seven were loaded into a pull-and-cut straw and then cryopreserved in liquid nitrogen. For thawing, the recovered embryos were treated with thawing solution mixed with 1 M sucrose (thawing solution 1) and 0.5 M sucrose (thawing solution 2) in plain medium for 1 and 3 minutes, respectively. The embryos were then twice incubated in plain medium for 5 minutes, followed by washing with blastocyst incubation medium, which was incubation 4 hours at 37℃ in 6% CO2.
5. Double fluorescence staining of blastocysts
In order to stain the TE cells, solution 1 was prepared by mixing 100 µg/mL of propidium iodide (P-4170, Sigma) and 1% Triton X-100 (T-93443, Sigma) in Ham's F-10 solution. Solution 2 was a mixture of 25 µg of bisbenzimide (B-2261, Sigma) in 99.9% ethanol, which was prepared to stain the ICM. In order to carry out double fluorescence staining, the blastocysts were stained in solution 1 for 10 seconds and transferred to solution 2 for 2 hours or more at 4℃. The stained blastocysts were washed with glycerol (G2025, Sigma) and put onto a slide glass for observation under a fluorescence microscope (BX50, Olympus, Japan) (Figure 2).
6. Statistical analysis
The survival rates of the blastocysts and the rates of formation of hatched blastocysts were expressed as percentages. The data were collected in Microsoft Office Excel 2007, and the chi-squared test was used to analyze discontinuous variables and standard errors as well as to compare the number of blastocysts between groups. A p-value of less than 0.05 was considered to indicate statistical significance.
Results
1. Effects of AS and AH on the development of blastocysts after V-T
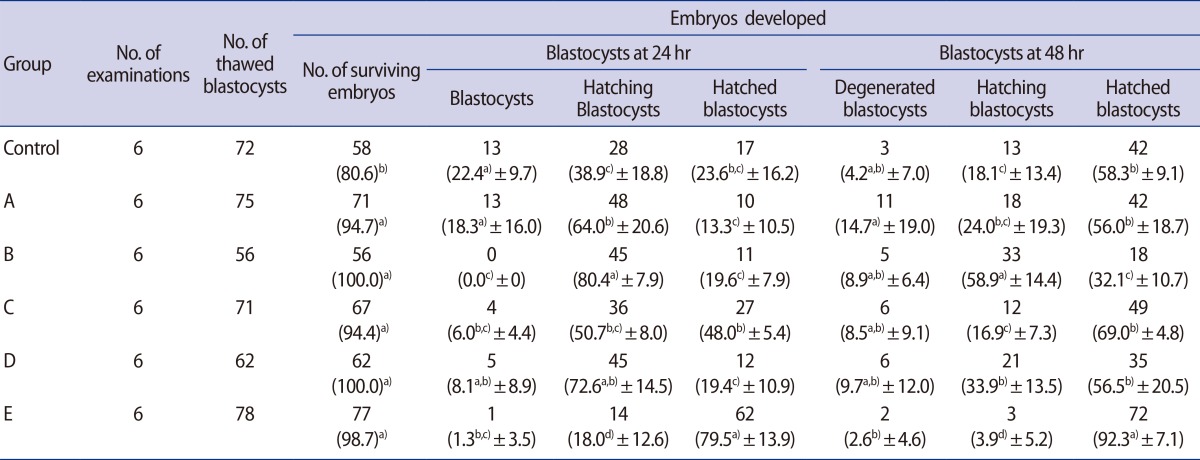
The survival rates of blastocysts in the cMAH and qLZT-AH groups, which were treated before vitrification and after thawing, as well as their developmental rate 48 hours after thawing, are presented in Table 1. The survival rates after thawing in the experimental groups (A, 94.7%; B, 100%; C, 94.4%; D, 100%; E, 98.7%) were significantly higher than in the control group (80.6%) (p<0.05). The rate of blastocyst formation during hatching was the highest in group B (AS/V-T/cMAH) at 24 and 48 hours after thawing. The rate of hatched blastocyst formation was significantly higher in group E (79.5%, 92.3%) than in the other groups (control, 23.6%:58.3%; A, 13.3%:56%; B, 19.6%:32.1%; C, 48%:69%; D, 19.4%:56.5%) at 24 and 48 hours after thawing (p<0.05).
2. Effects of AS and AH on the cell number of blastocysts after V-T
Table 2 summarizes the ratios of the cell number of the ICM to the total blastocyst and TE cell numbers in double fluorescence-stained hatching and hatched blastocysts after the cMAH and qLZT-AH procedures were carried out before vitrification, and after 48 hours of postcultivation. In the present experiment, embryos that were lost during the staining procedures, stained singly, or stained in an overlapped position with fuzzy borders were discarded, resulting in a total number of 258 embryos, instead of 358. The cell numbers in the ICM of the stained blastocysts were significantly higher in the control group (21.4±7.3), and group A (21.9±7.5), and group B (20.6±7.1) than in other groups (C, 17.1±5.0; D, 16.3±3.9; E, 16.5±5.2) (p<0.05). Additionally, the number of cells in the TE and the total cell number were higher in the control group (77.9±17.5:99.3±18.3) than in the other groups (A, 69.5±18.2:91.4±21.6; B, 72.2±17.5:92.8±18.8; C, 46.8±15.2:64±14.9; D, 59.8±13.9:76.2±15.9; E, 56±13.7:72.5±15.4). The ratio of the cells in the ICM to the total cell number was higher in group C (26.8±9.0) than in the other groups (control, 21.5±7.0; A, 24±7.1; B, 22.2±7.0; D, 21.4±4.6; E, 22.8±6.2).
Discussion
Unlike early blastocysts, expanded blastocysts have a large cavity filled with fluid, known as a blastocoel. For this reason, an additional dehydration procedure is needed to remove this fluid during thawing. Studies on AS and cooling velocity have been conducted to overcome this issue. Vanderzwalmen et al. [16,17] reported that AS after dehydration of the blastocoel using a glass micropipette smaller in diameter than the embryo increased the pregnancy rate after cryopreservation by 20%. They also reported that the pregnancy rates were increased by 38% and 19%, respectively, in experimental groups with and without the AH technique, which involves by partial zona dissection after AS with a glass micropipette that penetrates the blastocyst. Son et al. [18] reported that the survival and pregnancy rates were increased by 90% and 48%, respectively, by cryopreservation and thawing after partial zona dissection of the blastocyst using a 29-gauge needle in the 6 o'clock or 12 o'clock position of the ICM, followed by AS and AH. Additionally, Jo et al. [21] reported that the survival rates of cryopreserved blastocysts after AS were 98% and 92% with and without AH, respectively, while the survival rates of cryopreserved blastocysts without AS were 54% and 58% with and without AH, respectively. This led to the conclusion that AS is an efficient method for increasing the viability rate after the cryopreservation of mouse blastocysts, regardless of AH. In the present study, the survival rates of blastocysts that underwent AS and AH both before and after cryopreservation- and thawing were higher than the survival rates of the blastocysts in the control group AS and AH (Table 1). When compared to the effect of AS on the likelihood of pregnancy associated with cryopreserved and thawed embryos reported by Tucker et al. [22], Check et al. [23], and Tao and Tamis [15], the pregnancy rates in the groups with and without AH were 28% vs. 15%, 30.4% vs. 15.2%, and 24% vs. 0%, respectively. These results mean that AH may have a positive impact on the pregnancy rate. Jo et al. [21] reported that the hatching rates associated with AH using the PZD method were 96% and 34% in the groups with and without AS, respectively. The hatching rates of the groups with and without AH after AS were 100% and 41%, respectively. A high hatching rate was found after AH in cryopreserved and thawed blastocyst embryos (p<0.01), and they concluded that AH was an effective method for improving the hatching rate. Cieslak et al. [24] reported that a pregnancy rate of 42.0% was obtained in a group that underwent three-dimensional partial zona dissection, using a micropipette rather than strong acid for dissecting the ZP in preimplantation genetic diagnosis and AH, which was then rotated for another round of dissection, compared to a rate of 33.3% in the control group. Additionally, Park et al. [25] showed that the rate of fully hatched embryos cultivated for 72 hours after AS was 9.0% in the group in which cMAH was performed, compared to 0% in the group in which single mechanical AH. The present experiment was largely based on the report of Khalifa et al. [26], who found a high incidence of hatched blastocyte embryo formation using quarter-LAH (qLAH) and qLZ-AH, which are helpful techniques for encouraging blastocysts to hatch because they eliminate the rather large area of the ZP. The rates of hatched blastocyst formation were 49.1% and 40.4% in the qLAH and qLZT-AH groups, respectively, which were significantly higher than in the groups which a smaller area of the ZP was eliminated. Their study was referenced by Park et al. [25], who found a significantly higher rate of formation of hatched blastocysts after 72 hours of culture in the qLAH and qLZT-AH groups than in the control groups. In the present study, the rate of hatched blastocyst embryos was consistently the highest in the AS/qLZT-AH/V-T group (Table 1).
According to Thouas et al. [27] the cell numbers of the ICM and TE and the total cell numbers of 2-cell mouse embryos were 21±2, 55±4, and 75.3±3, respectively, after 72 hours of cultivation. The cell number in the ICM of the control group was similar to that found in this study (21.4±7.3), however, some discrepancies were present in the cell number of the TE (77.9±17.5) and in the total cell number (99.3±18.3) (Table 2). It has been reported that faster embryonic development leads to the formation more cells and a higher ratio of the ICM to total cells [28]. However, Park et al. [25] reported that the formation rate of the hatched blastocyst embryos in the qLZT-AH group was higher than what was observed in the cMAH group (p<0.05). They argued that the improved hatching results were not due to the large cell number because the number of cells in the TE was low. In the present study, the formation rate of hatched blastocyst embryos in the AS/qLZT-AH/V-T group, which underwent cryopreservation after AS and qLZT-AH, was the highest among all experimental groups. However, in this group, the cell number of the ICM and TE and the total cell number were low, similar to the results of Park et al. [25] (Table 2). Lane et al. [29] reported that the cell number of the TE was similar to that of the blastocysts regardless of the implantation technique, as well as no correlation between the number of cells in the TE and the implantation rate in mouse blastocyst embryos. Richter et al. [30] reported that the implantation rate did not vary with the size of the blastocyst, the number of cells in the TE, or the size and shape of ICM, and only varied according to the external shape of the embryo. In the present study, the survival rate of blastocyst embryos after thawing was significantly different depending on whether AS before cryopreservation, but the hatching rate was similar.
The highest formation rate of expanded blastocysts was achieved by performing the AS and qLZT-AH procedures before vitrification, followed by 48 hours of cultivation. The next highest formation rates were observed after the cMAH procedure, in AS and AH were performed with a micropipette, and the qLZT-AH procedure, which used a laser before V-T. The cell number of in the ICM and TE and the total cell count were significantly higher in the control group, in which AS and AH were not performed, than in other groups. With regard to the above results, AS and qLZT-AH procedures performed before cryopreservation are known to have beneficial effects on embryonic development. With this in mind, further studies are needed regarding the timing and methodologies of AH in embryo transfer procedures involving cryopreservation and thawing.