Introduction
Triploidy is one of the most common chromosome abnormalities in human gestation, as it is present in 1%-2% of pregnancies [1]. However, triploid pregnancies rarely come to term, and the very diverse phenotypes seen, are all grossly abnormal [1,2]. Triploidy may originate from either digynic or diandric fertilizations, involving either a diploid oocyte or dispermic fertilization of an oocyte [3,4,5].
Triploidy is also found in vitro and occurs in 4%-7% of in vitro fertilized [4,6,7] and 6% of intracytoplasmic sperm-injected (ICSI) zygotes [4]. The vast majority of triploid embryos are recognized before transfer to the woman, by the appearance of 3 pronuclei (3PN) in the zygote instead of the normal 2. Otherwise, 3PN embryos and embryos with two pronuclei (2PN) have the same morphological appearance [8]. Analyses of 3PN zygotes after in vitro fertilization (IVF) have indicated that at least 86% originate from dispermic fertilization of the oocyte [9]. Since only one spermatozoon is injected into the oocyte at ICSI fertilization, the extra pronucleus in 3PN ICSI embryos is most likely of maternal origin. As the centriole is paternally inherited-supplied as an organelle by the sperm-3PN IVF and ICSI zygotes tend to have different numbers of centrioles [10,11].
Although it has been demonstrated that 3PN zygotes have 3 haploid sets of chromosomes [3,10,12,13,14,15], several studies of cleavage stage embryos arising from 3PN zygotes have revealed karyotypes that are triploid, diploid, or severely abnormal including mosaicism [4,6,8,12,13,16,17,18]. Conclusions from these studies are, however, scarce as only a few nuclei from each embryo have been analyzed, or a low number of chromosomes have been analyzed from each nucleus [4,6,8,12,13,16,17,18].
Only a few studies have analyzed any potential differences in karyotypes between 3PN IVF and 3PN ICSI embryos [4,6,12,18], and only two studies stratified cases according to the embryonic age at which the embryos were analyzed [19,20].
Therefore, further analyses of the karyotypes in 3PN IVF and 3PN ICSI embryos could provide essential insight into early embryo development, the role of centrioles in human embryogenesis, and basic genetics.
In the present study, 163 nuclei collected from 37 3PN embryos were analyzed for ploidy using fluorescence in situ hybridization (FISH) probes for 8 different chromosomes. The results were analyzed with a focus on the fertilization method (IVF or ICSI) and the embryonic age.
Methods
1. Ethical approval
Between May 1, 2010, and June 30, 2011, all patients undergoing treatment at the Fertility Clinic of Braedstrup/Horsens Hospital were asked for permission to include their 3PN zygotes in the study.
Oral and written informed consent was obtained from all participants before inclusion. The Regional Committees on Biomedical Research Ethics in Southern Denmark and the Danish Data Protection Agency approved the study.
2. Participants and IVF
Patients underwent treatment according to the guidelines in the clinic; that is, standard IVF was performed in all cases unless there were (1) less than 1 million spermatozoa/mL after preparation, (2) low spermatozoa motility, or (3) a history of IVF failure. In these instances, standard ICSI procedures were followed.
3. Embryo culture
Immediately after sperm addition or injection, fertilized embryos were cultured under oil at 37℃, 6% CO, and 5% O2 in a sequential culture medium (Cook Sydney IVF Fertilization/Cleavage/Blastocyst Medium, Cook, Sydney, Australia) changed on days 3 and 5. At 18-20 hours after injection or addition of sperm, the oocytes were checked for fertilization via the counting of pronuclei. If a 3PN zygote was observed and the patient had consented, the zygote was included in the study.
4. Embryo fixation
Artificial arrest of the developing 3PN embryos was performed at different points in time between day 0 and day 5. During the fixation procedure, embryos were observed using an inverted microscope.
Embryos were removed from the culture medium and incubated in a drop of Tyrode's acid in order to remove the zona pellucida. Just before complete disintegration of the zona pellucida, the embryos were transferred to a drop of phosphate buffered saline (PBS). The embryos were then transferred to a poly-L-lysine slide (Menzel-Glaser, Germany) along with 0.01-mol/L hydrogen chloride (HCl) in 0.1% polysorbate 20 (Tween 20). At this point, the cell membrane and the cytoplasm began to disintegrate, leaving the nuclei clearly visible and fixed (after a few seconds to several minutes). The location of the nuclei was registered and marked by a diamond pen. Slides were air-dried followed by washing in PBS and dehydrated using an ascending ethanol series. The slides were stored at -80℃ until the hybridization protocol was performed. For each embryo, the number of cells observed before fixation and the number of nuclei observed after fixation were noted.
5. FISH
FISH of the fixed nuclei was performed using four sequential hybridizations. Centromeric probes for chromosomes 6 and 11 (first hybridization), 8 and 17 (second hybridization), 10 and 18 (third hybridization), and X and Y (fourth hybridization) were used-all commercially available from Cytocell, Cambridge, United Kingdom.
The hybridization protocol, in brief, was as follows: Slides were incubated with pepsin (100 µg/mL) for 10 minutes at 37℃, rinsed in bi-distilled water and PBS, and fixed for 5 minutes in 1% paraformaldehyde in PBS at room temperature. The slides were then rinsed in PBS and bi-distilled water and dehydrated using an ascending ethanol series. The probes were added to the slide under a coverslip. The probe and the nuclear DNA were denatured at 75℃ for 3 minutes and left to hybridize in a humidity chamber at 37℃ for 1 hour. To remove an unhybridized probe, the slides were washed in 50% formamide in saline-sodium citrate buffer (SSC) at 42℃ for 5 minutes, followed by washing in SSC and 0.05% polysorbate 20 (Tween 20) at room temperature for 5 minutes. The slides were dehydrated and mounted in Vectashield (Vector Laboratories, USA) antifade medium containing 1 ng/mL 4,6-diamidine-2-phenylindole to counterstain the nuclei and examined.
In order to perform rehybridization, the slides were washed twice in SSC and 0.05% polysorbate 20 (Tween 20) at room temperature for 5 minutes, followed by 2 minutes of washing in PBS and then dehydrated through an ascending ethanol series, after which the denaturation and hybridization could proceed as mentioned above.
The nuclei were analyzed using an epifluorescence microscope (Leica DM6000B) and the images of every nucleus were recorded vertically (z-stacking) with an average distance between images of 0.8 µm using a digital monochrome CCD camera (DFC360FX) operated by the Leica Application Suite Advanced Fluorescence software.
A microscope slide containing lymphocyte nuclei from a male with a normal karyotype was also processed during each of the FISH rounds, allowing us to monitor the efficiency of probe hybridization.
Analyses of the FISH signals were carried out using direct microscopy by a single observer (Obs. 1) without any knowledge of the fertilization method. Reproducibility was tested by a reanalysis of one image created by the merging of the z-stacked images of the nuclei into one. This reanalysis was performed by Obs. 1 (Obs. 1b) and by a second observer (Obs. 2), both blinded with respect to the fertilization method and previous results.
The following scoring criteria were used to assess the chromosome numbers: In cases where there was doubt if a fluorescence dot was a valid signal or an artifact, the number of signals for that specific chromosome was classified as unknown. Two signals represented two chromosomes if they were a minimum of a signal's width apart.
6. Data classification
Embryonic age at fixation was classified as follows: 0-2 days and 3-5 days, respectively. For each nucleus, the number of signals from one chromosome was classified into three groups: Chromosome count <3 (if the number of signals was smaller than 3), chromosome count =3 (if the number of signals was equal to 3), and chromosome count >3 (if the number of signals was larger than 3).
7. Statistical analyses
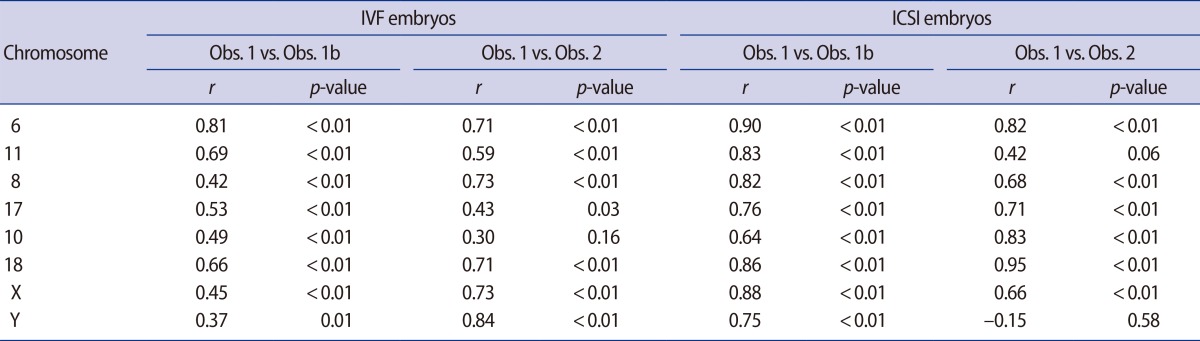
A two-tailed Mann-Whitney test was used to test the hypothesis of no difference between categorical baseline data, and a bootstrap likelihood ratio test (10,000 bootstrap samples) was applied for testing the possible differences in the number of nuclei per embryo analyzed between ICSI and IVF embryos [21]. Reproducibility (Obs. 1 vs. Obs. 1b, and Obs. 1 vs. Obs. 2) was tested using the non-parametric Spearman's correlation to compute the correlation coefficient (r) between the counts for each chromosome reanalyzed.
The probabilities of occurrence of chromosome count <3 and chromosome count >3 were studied using a multivariate Bernoulli (i.e., binomial with size 1) logistic mixed model [22] with three Gaussian random components designed to account for the structural correlation of the experiment. The models included fixed effects representing the fertilization method, the embryonic age, and the chromosome observed. The three nested random components used were as follows: (1) Identity, taking the same value for all the observations arising from the same embryo; (2) Nucleus, taking the same value for each observation arising from the same embryo and the same observed nucleus, and (3) Chromosome identity, taking the same value for each observation from the same embryo, the same observed nucleus, and the same chromosome. In these analyses, the numbers of signals from chromosomes X and Y were combined. The model allowed the use of the three variance components referred above to be different for the observation submitted to different fertilization methods. Note that, due to the basic properties of the Bernoulli logistic model used and because the random components above are nested (in the order given), the probability of two observations arising from the same nucleus and the same individual but related to different chromosomes present the same number of signals increases linearly with the total variance, i.e., the sum of the variances of the three random components [21,23,24]. This fact will be explored to characterize the degree of coordination of the number of signals for different chromosomes (i.e., the closeness to euploidy).
The statistical significance of the fixed effects (fertilization method, embryonic age, and chromosome-count) were tested using a parametric bootstrap version of the likelihood ratio test with 10,000 bootstrap samples (here we used a bootstrap version of the likelihood ratio test because the asymptotic distribution of the statistic of the likelihood ratio test under binomial logistic mixed models is not known).
Results
Sixty-two 3PN embryos were fixated (51 IVF and 11 ICSI). FISH failed in 16 3PN embryos (14 IVF and 2 ICSI). 3PN embryos where only three or fewer FISH probes could be analyzed in all nuclei were excluded from the analysis. In a total of 38 (31 IVF and 7 ICSI) 3PN embryos corresponding to 163 (131 IVF and 32 ICSI) nuclei, the number of signals for 8 different chromosomes were estimated. Baseline data for the embryos analyzed are presented in Table 1. The number of signals was obtained for 993 chromosomes (773 IVF and 220 ICSI), and representative data of the FISH results are shown in Figure 1.
There was a high correlation between the initial chromosome count and the chromosome count made by Obs. 1b and Obs. 2 (Table 2). The initial chromosome counts made by Obs. 1 were used for further analyses.
Chromosome count <3 was found for 69.6% (691/993) of the chromosomes, chromosome count =3 was found for 22.7% (225/993) of the chromosomes, and chromosome count >3 was found for 7.8% (77/993) of the chromosomes.
The probability of occurrence of chromosome count <3 and the probability of occurrence of chromosome count >3 were found not to depend on embryonic age nor on the specific chromosome analyzed (p=0.21 and p=0.31, respectively). On the other hand, the probability of chromosome count <3 and chromosome count >3 both depended significantly on the fertilization method (IVF or ICSI). Indeed, the difference between the probability of occurrence of chromosome count <3 and the probability of occurrence of chromosome count =3 were significantly greater for nuclei from 3PN ICSI than for nuclei from 3PN IVF embryos (odds ratio [OR], 5.28; 95% confidence interval [CI], 1.13-24.75, p<0.001) Likewise, the difference between the probability of occurrence of chromosome count >3 and the probability of occurrence of chromosome count =3 were significantly greater for nuclei from 3PN ICSI than for nuclei from 3PN IVF embryos (OR, 4.35; 95% CI, 1.09-17.38; p<0.001).
The probabilities of two observations arising from the same nucleus and the same individual but related to different chromosomes presenting the same number of signals were significantly higher in 3PN ICSI than 3PN IVF fertilized embryos (indeed, the total variances, which increase linearly with the total variance, were 14.47±3.81 and 12.87±1.69 for ICSI and IVF, respectively, p<0.001) for chromosomes with chromosome count <3. Similar results were found for chromosomes with chromosome count >3 (the total variances were 4.123±2.030 and 2.87±1.69 for ICSI and IVF, respectively, p=0.002). This indicated a higher degree of coordination of the number of signals for the various chromosomes analyzed in 3PN ICSI embryos than 3PN IVF embryos.
Discussion
In our cohort, only the fertilization method correlated to differences in chromosome count. In both 3PN IVF and 3PN ICSI embryos, there was a tendency for a reduction in the number of chromosomes from the assumed number of 3 copies of each chromosome in the 3PN zygote. In 3PN ICSI embryos, the probability of a chromosome count below or above 3 was significantly higher than in 3PN IVF embryos. A higher degree of coordination of the number of signals for different chromosomes were found in 3PN ICSI embryos compared to 3PN IVF embryos, indicating that the karyotypes of 3PN ICSI embryos were closer to euploid than those of 3PN IVF embryos.
1. Limitations to the study
FISH analysis for a limited number of chromosomes was used to estimate the chromosomal status of the 3PN embryos. Therefore, we were not able evaluate untested chromosomes. Likewise, FISH does not allow for detection of partial deletions or duplications of chromosomes since the probe hybridizes to a specific locus or the centromere, and hence only provides information about that segment of the chromosome [27]. Another technical limitation of FISH is that errors in chromosome count can occur due to loss or damage of nuclear material; split, diffused, or overlapping signals; hybridization failure; or probe inefficiency. As FISH analyses have approximately 95% accuracy per probe, using a multiprobe panel on one nucleus from a 3PN embryo, potentially with several signals per probe, renders the risk of misdiagnosis significant [28,29]. Nevertheless, FISH is as reliable technique for ploidy determination for research purposes [29]. Moreover, we controlled the chromosome count by evaluating the reproducibility and found high intra- and inter-observer correlations. FISH failed in some embryos and as only high quality FISH results were analyzed, a smaller cohort was available for further analyses. Despite the relatively small cohort analyzed, significant differences were found. Our data does not, however, allow for exclusion of variables other than the fertilization method (for example maternal/paternal age), even though they also could impact the karyotype of the embryos.
2. Chromosomal makeup of 3PN embryos
Although it has been demonstrated that 3PN zygotes generally have 3 haploid sets of chromosomes, several studies of 3PN embryos have revealed triploid, diploid, or severely abnormal karyotypes [3,4,6,8,10,12,13,14,15,16,17,18,19,30]. However, studies reporting mosaic chromosome imbalances in normal 2PN in vitro fertilized embryos showed that chromosome instability also occurs during normal human embryogenesis [31,32,33,34,35,36,37,38,39,40]. This instability could be a result of the inactive embryonic genome rendering normal mechanisms for induction of apoptosis in chromosomal abnormal cells impossible [33,34,38]. This is supported by reports suggesting a decrease in the proportion of aneuploid cells at the blastocyst stage [33,35,38].
The aneuploidy and mosaicism previously reported in 3PN embryos [3,4,6,8,10,12,13,14,15,16,17,18,19,30] could, therefore, be a "normal" developmental event with an etiology as mentioned above and not a consequence of the 3PN status.
If the aneuploidy and mosaicism found in 3PN embryos were the effect of a "normal" developmental event alone, there would presumably be no difference in the degree of coordination of the number of signals for different chromosomes in 3PN ICSI and IVF embryos.
However, 3PN ICSI nuclei displayed a higher degree of coordination between numbers of signals for different chromosomes than 3PN IVF nuclei. A possible explanation might be different numbers of centrioles in the 3PN zygote.
In the human zygote the first mitotic division is controlled by the paternally inherited centriole [10,11]. Fertilization with two spermatozoa introduces 2 centrioles into the oocyte. Abnormal cleavage directly into 3 cells, has been reported in cells with extra centrioles [5,7,10,11,41]. The vast majority of 3PN IVF zygotes are of dispermic origin and thus more likely to have extra centrioles [42]. Concordantly, we have previously observed that significantly more 3PN IVF zygotes cleaved early into 3 cells-defined as a duration of the 2-cell stage of less than 1 hour-than either 3PN ICSI or 2PN IVF zygotes [43]. If, in fact, the cells showing "early cleavage into 3 cells" actually undergo direct cleavage into 3 cells, then the 3PN IVF embryos would be expected to display a reduction in the number of chromosomes per cell, since the triploid genome - after duplication and syngamy of the pronuclei-is divided into 3 cells instead of the normal 2. This could explain why 3PN IVF nuclei in this study tended to have a chromosome count <3.
The different cleavage patterns observed for 3PN IVF and 3PN ICSI embryos do, however, not explain why 3PN ICSI nuclei were more likely than the 3PN IVF nuclei to display a chromosome count below or above 3. The 3PN ICSI nuclei displayed a higher coordination of the number of signals for different chromosomes than the 3PN IVF nuclei, which could indicate that 3PN ICSI nuclei possess a mechanism of euploid ploidy reduction (removal of haploid set(s) of chromosomes). As Grau et al. [19] have found that self-correction in 3PN ICSI embryos apparently takes place before day 2 of embryo development and hence, before embryonic genome activation, we speculate that the self-correction is independent of genomic content but caused by an insufficient number of centrioles relative to the number of pronuclei. Such an insufficiency could hamper the pronuclear apposition of all pronuclei after fertilization.
No studies have analyzed the pronuclear apposition in detail in humans. However, studies of other species have shown that pronuclear apposition is dependent on the ability of the sperm asters to both push the male pronucleus further into the cytoplasm, and to reach the female pronucleus allowing it to be moved toward the male pronucleus [44,45].
We, therefore, speculate that this apparent ability to self-correct or maintain polyploidy in 3PN ICSI embryos, is caused by an insufficient number of sperm asters relative to the number of pronuclei, speculating that one sperm aster may not always form adequate numbers of microtubules to allow 3 pronuclei to assemble. In that case, this self-correction of the polyploidy would be less likely in the 3PN IVF embryos.
Our results show that the fertilization method (IVF or ICSI) impacts the karyotype of the 3PN embryos. The chromosome count in the ICSI nuclei tended to differ more from 3 and were closer to euploidy than that in the 3PN IVF nuclei. This could indicate that the number of centrioles in the 3PN zygote influences the karyotype.