Introduction
Autophagy is a self-consuming subcellular process that occurs widely in various tissues. While it takes place at a basal level in all tissues, the consequences of autophagy deficiency vary significantly depending on the tissue type. In the mouse uterus, autophagy levels differ between epithelial and mesenchymal tissues [1]. We have previously demonstrated that the uterine mesenchyme, including the stroma, myometrium, and endothelial smooth muscle cells, is a site of active autophagic turnover in mice [2]. Deletion of the upstream autophagy factor, autophagy-related gene 7 (Atg7), which encodes an E1 ubiquitin-activating enzyme, blocks autophagic flux, and is a widely used model system to investigate the role of autophagy [3]. In the mouse uterus, cross-breeding Atg7 floxed mice [3] with anti-M├╝llerian hormone receptor type 2 receptor-Cre (Amhr2-Cre) knock-in mice [4] produces uterine mesenchyme-specific deletion of Atg7, while its expression remains intact in the epithelium. Various vasoactive factors accumulate in Atg7f/f;Amhr2-Cre female mice, leading to hyperpermeability and fluid retention in their uteri [2]. This phenotype is most prominent when steroid hormones are removed via ovariectomy. Random cycling mice also gradually develop similar phenotypes at a slower pace; thus, this phenotype is much more visible in older female mice [2]. The accumulation of vasoactive factors does not interfere with pregnancy, and Atg7f/f;Amhr2-Cre mice can be used at breeders.
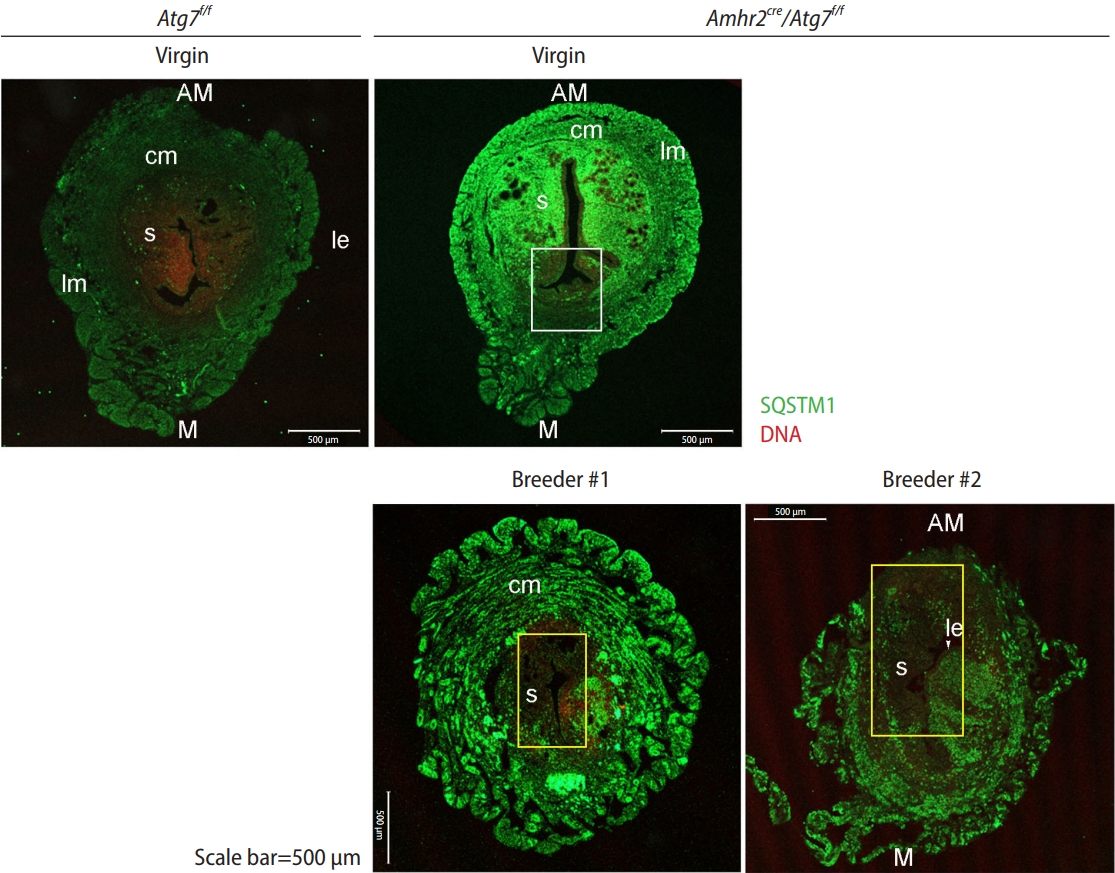
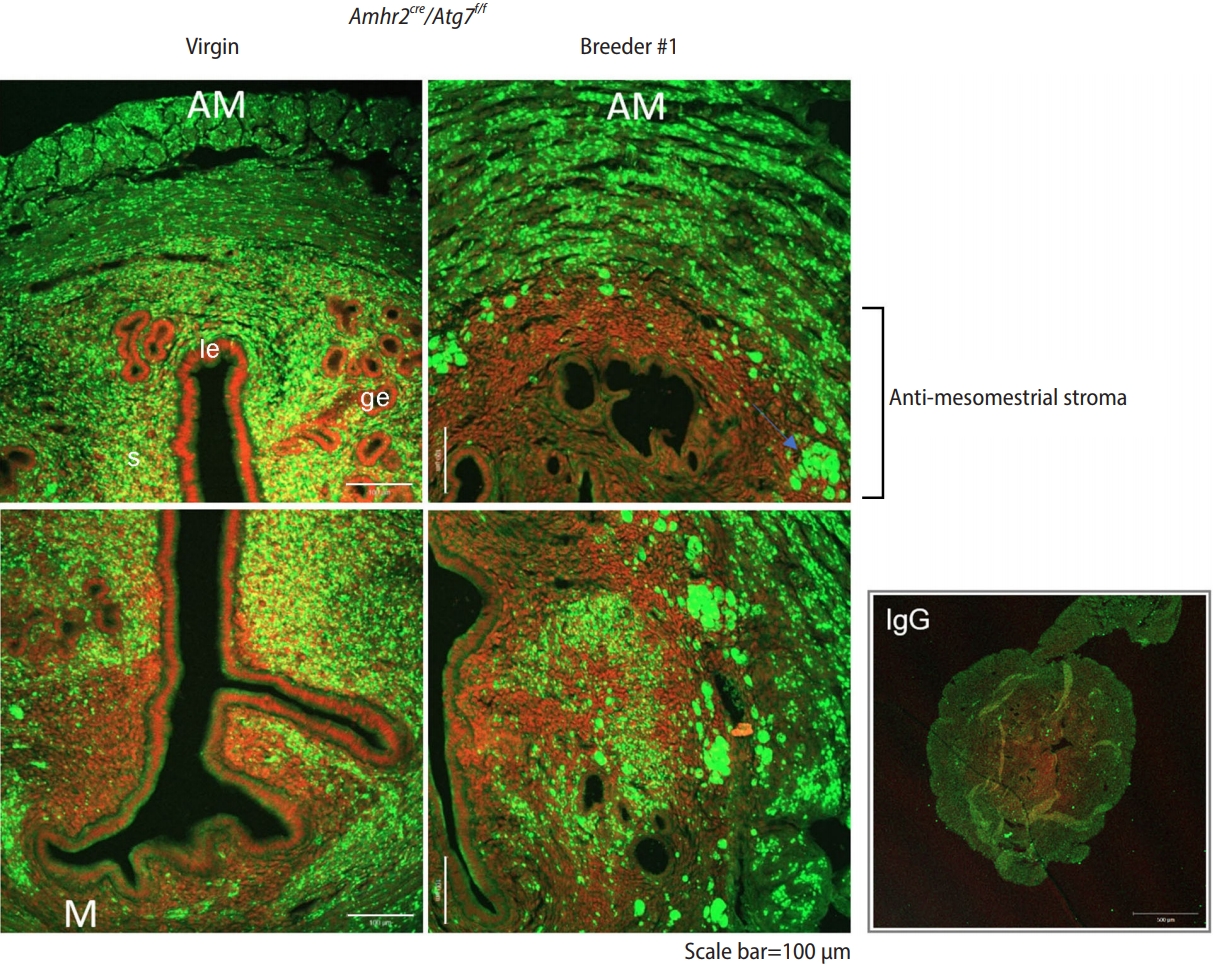
Amhr2-Cre mice have been extensively used to study gene function in the mouse uterus [5-7]. Amh2-Cre removes a floxed gene in the M├╝llerian duct mesenchyme-derived cells in the uterus, but a subset of stromal cells in the mesometrial side (the future site of placental attachment) does not express Amhr2 [8], retaining the floxed gene in this sub-region of the uterine stroma. This area is proportionally quite narrow compared with the Amhr2-active region. In Atg7f/f;Amhr2-Cre uteri, the differing status of gene deletion was visualized using sequestosome 1 (SQSTM1), as an autophagic deficit leads to SQSTM1 buildup in the tissue. As shown in Figure 1, the Atg7-deleted stromal region in the anti-mesometrial side was distinctively marked with anti-SQSTM1 immunofluorescence staining, whereas the myometrial stromal subregion of intact Atg7 showed no SQSTM1 signal (white rectangle) (Figure 1, virgin Atg7f/f;Amhr2-Cre). Thus, this subregion in the mesometrial mesenchyme of the Atg7f/f;Amhr2-Cre uterus retains autophagy and always shows similar patterns of SQSTM1 signal regardless of age [2].
The uterus has a regenerative capacity essential for maintaining homeostasis during repeated reproductive cycles and pregnancy [9]. The mouse endometrium has been shown to retain certain stem cell populations both in the epithelium and stroma [10]. Amhr2-Cre mice crossbred with a reporter gene were previously used to examine how uterine cells regenerate during repeated estrous cycles or postpartum [11]. Stem cells in the epithelial and stromal compartments retain their original fates during repeated estrous cycles, whereas postpartum endometrial regeneration involves a transition from stromal to epithelial cells [11]. This study demonstrated that postpartum uterine regeneration differs from cycle-dependent regeneration during homeotic maintenance of the uterus.
In this study, we examined whether the composition of stromal cells changed after repeated parturition. Following the pattern of SQSTM1 accumulation, we discovered that Atg7-deficient stromal cells were in part replaced by Atg7-intact cells in the uteri of Atg7f/f;Amhr2-Cre breeders, and the phenotype caused by the autophagic deficit seemed to have diminished in them as well.
Methods
1. Mice
Amhr2-Cre knock-in mice [4] and Atg7 floxed (Atg7f/f) mice [3] were obtained from Dr. Richard Behringer and the RIKEN BioResource Center (Ibaraki, Japan), respectively. Amhr2-Cre;Atg7f/f mice were crossbred with Institute of Cancer Research mice for more than six generations to obtain mice with better reproductive performance. Amhr2-Cre;Atg7f/f female mice exhibited normal fertility phenotypes equivalent to those of Atg7f/f female mice with respect to the number of deliveries and pups per litter during the breeding period [2]. To compare the status of uterine autophagy between virgins and breeders, Amhr2-Cre;Atg7f/f female mice were separated into virgin (10 to 12 months old) and breeder groups. Virgin mice were not used in breeding and were kept separately from male mice until they reached 10 to 12 months. Breeders, from the ages of 6 to 7 weeks until they reached 10 to 12 months of age, were continuously bred with a stud Atg7f/f male. All mice were maintained according to the policies of the Konkuk University International Animal Care and Use Committee (IACUC approval number KU19078).
2. Immunofluorescence staining
Uteri were collected from female mice with the indicated genotypes, cut into 3 to 5 mm pieces, and fixed in 4% paraformaldehyde (PFA) in phosphate-buffered saline (PBS) overnight at 4 ┬░C. The uterine pieces were washed with PBS and embedded in sucrose solution for several hours until the pieces were completely submerged in the solution. They were flash-frozen in optimal cutting temperature compound and subjected to cryosection (12 ╬╝m thickness). The uterine sections were then fixed in 4% PFA-PBS for 20 minutes, washed, and permeabilized with 0.1% Tween-20 in PBS for 20 minutes. The specimens were blocked with 2% bovine serum albumin (BSA) in PBS for 1 hour at 25 ┬░C and then incubated with anti-SQSTM1 antibody (Cell Signaling Technology) in 2% BSA-PBS for 2 hours at 25 ┬░C [2]. After washing, the sections were incubated with Alexa Fluor 488-conjugated secondary antibodies (Thermo Fisher Scientific) for 40 minutes. The sections were counterstained with TOPRO-3-iodide (Thermo Fisher Scientific). The mounted slides were observed by live confocal imaging using a Zeiss LMS900 confocal microscope.
3. Western blotting
Uteri were collected from each mouse, cut into 5-mm pieces, and placed in lysis buffer containing 10 mM Tris (Thermo Fisher Scientific; pH 7.2), 150 mM NaCl (Thermo Fisher Scientific), 0.1% Triton X-100 (Sigma Aldrich), 5 mM ethylene diamine tetraacetic acid (Thermo Fisher Scientific), 1% sodium dodecyl sulfate (SDS, Thermo Fisher Scientific), 1 mM dithiothreitol (Sigma Aldrich), 1 mM phenylmethylsulfonyl fluoride (MP Biomedicals), and 1├Ś protease inhibitor (Roche). Each sample was homogenized and centrifuged at 15,928 ├Śg for 20 minutes at 4 ┬░C. Protein concentrations were determined by the bicinchoninic acid protein assay (Thermo Fisher Scientific). Lysates (10 ╬╝g) were loaded onto SDS-polyacrylamide gels and transferred onto polyvinylidene difluoride membranes (Sigma Aldrich). The following reagents were used to prepare SDS-polyacrylamide gels: 30% acrylamide mix (Bio-Rad), ammonium persulfate (Sigma Aldrich), TEMED (Sigma Aldrich), 1.5 M Tris (Biosesang; pH 8.8), and 0.5 M Tris (Biosesang; pH 6.8). Chemiluminescent signals were quantified using the ImageQuant LAS4000 system (GE Healthcare) and Multi Gauge software (GE Healthcare). The signal intensity was normalized to the ╬▓-tubulin signal. Data are shown as mean┬▒standard error of the mean of six independent biological samples. The primary antibodies used were anti-SQSTM1, anti-╬▓-catenin (anti-CTNNB1, Cell Signaling Technology), anti-endothelin 1 (anti-EDN1, Biorbyt), anti-nNOS (Cell Signaling Technology), anti-vascular endothelial growth factor A (anti-VEGFA, Abcam), and anti-╬▓-tubulin (Abcam) [2].
Results
1. Changes in the pattern of SQSTM1 accumulation in breeder Atg7d/d/Amhr2-Cre uteri
Autophagic defects are generally visualized by examining SQSTM1 accumulation [1] because it is highly upregulated when autophagic flux is blocked [12]. In Atg7f/f (wild-type) virgin uteri, autophagy is generally active, and there was no visible accumulation of SQSTM1 (Figure 1, virgin Atg7f/f), which was consistent with the results of our previous study [2]. In Atg7f/f;Amhr2-Cre random-cycling virgin mice (>11-month-old mice), heavy SQSTM1 buildup was observed in the stroma and myometrium, but not in the luminal and glandular epithelial cells. In addition, the white rectangle marks the small mesenchymal region on the mesometrial side without the SQSTM1 signal, indicating that this area retains Atg7 and intact autophagy. The bottom panel shows uteri from two Atg7f/f;Amhr2-Cre breeders (n=2). In Atg7f/f;Amhr2-Cre breeders, the myometrium was more expanded than in virgin Atg7f/f;Amhr2-Cre mice. Yellow rectangles indicate areas with no SQSTM1 accumulation (Figure 1). In both breeders, the yellow rectangle encompasses the stromal region on the anti-mesometrial and mesometrial sides. The difference in SQSTM1 buildup between Atg7f/f;Amhr2-Cre virgin and breeder mice is clearly visible in Figure 2. The anti-mesometrial side with the supposed Atg7 deletion showed no SQSTM1 accumulation in breeder #1. These results suggest that uterine regeneration after repeated parturition results in the loss of subsets of Atg7-deleted stromal cells.
2. Functional restoration of autophagy in the uteri of breeder Atg7f/f;Amhr2-Cre mice
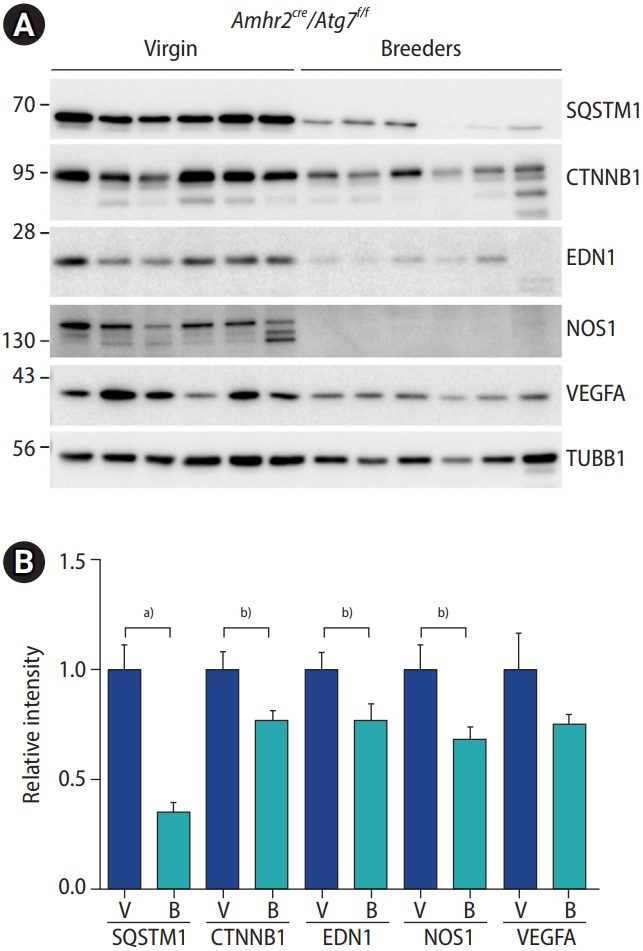
Next, we examined whether the functional aspects of autophagy were restored in the uteri of Atg7f/f and Amhr2-Cre breeders. As previously reported, in Atg7f/f;Amhr2-Cre mice, high levels of several vasoactive factors, including EDN1, nitric oxide synthase 1 (NOS1), VEGFA, and CTNNB1, were observed in their uteri compared to those in wild-type mice. This phenotype was visible in both random cycling and ovariectomized Atg7f/f;Amhr2-Cre uteri [2]. We examined the accumulation of the aforementioned vasoactive factors using Western blotting (Figure 3). Compared to virgin uteri (n=6), the uteri of breeders (n=6) showed significantly reduced accumulation of SQSTM1, CTNNB1, EDN1, NOS1, and VEGFA. These results suggest that postpartum uterine regeneration compensated for autophagy-deficient cells with autophagy-intact cells in the uterine mesenchyme of Atg7f/f;Amhr2-Cre mice.
Discussion
The uterine mesenchyme is a site of dynamic autophagy, which turns over vasoactive factors during repeated reproductive cycles. When mesenchymal autophagy is blocked by Atg7 deletion, the uterus exhibits hyperpermeability owing to an overload of vasoactive factors [2]. In the present study, we provide evidence that the mesenchymal cell population undergoes a turnover during repeated breeding in Atg7f/f;Amhr2-Cre mice. This observation was possible because of the unique expression profile of Amhr2, which drives Cre expression in this model. Amhr2 is widely expressed in the M├╝llerian duct mesenchyme during development, except for a small subset of mesenchymal cells in the mesometrial side where the broad ligament inserts [11]. When Atg7 is deleted in Amhr2-Cre mice, a subset of mesometrial mesenchymal cells retains the Atg7 gene; thus, the autophagic process is normally preserved in this region [2]. Because SQSTM1 accumulation is only observed in autophagy-deficient cells in the uterus, we were able to follow the potential cell mixing. As shown in Figures 1 and 2, autophagy-deficient mesenchymal cells appeared to have infiltrated the anti-mesometrial side in breeder Atg7f/f;Amhr2-Cre mice (Figures 1 and 2), whereas the uterus of virgin Atg7f/f;Amhr2-Cre mice showed autophagy-deficient mesenchymal cells on the anti-mesometrial side. Cell mixing is possible in the uteri of breeders as the uterus undergoes massive regeneration after parturition in mice [13].
A previous study using the same Amhr2-Cre knock-in mice for the removal of the beta-catenin gene (Ctnnb) in the uterine mesenchyme showed that fat lumps protruded from the anti-mesometrial side of the uterine horns in gene-deleted uteri [14]. It has been suggested that there is a progressive switch in fate from smooth muscle (composing the myometrium) to fat, both of which are of mesenchymal origin [14]. This fate switch was not evident in the mesometrial side where Amhr2-Cre expression was absent. Thus, the Amhr2-Cre knock-in model is useful for delineating the distinct roles of mesenchymal cells in the mesometrial and anti-mesometrial sides of the uterus. In our study, autophagy-deficient mesenchymal cells seem to have been overridden in part by autophagy-intact cells on the mesometrial side (Figures 1 and 2). This change in cell composition further influenced the functional aspect, reducing the burden of autophagic deficit in the whole uterus; that is, a reduction in certain vasoactive factor levels was noted in Atg7f/f;Amhr2-Cre breeders (Figure 3).
The pregnant uterus undergoes massive remodeling postpartum, a process termed uterine involution [15]. In this event of massive tissue remodeling, cell mixing and replenishment are likely to occur. The process involves cell repair by apoptosis, autophagy, extracellular matrix degradation, and proteinase activation, and is believed to involve the activation of certain stem cells [13,15,16]. The label-retaining cell approach has been used to identify the location of endometrial stem cells (ESCs) in the mouse uterus [17]. This approach, following up cells with long-term retention of a DNA synthesis label, identified potential sites of ESCs in the postpartum uterus, such as the subluminal epithelial stroma, near blood vessels, and near the endometrial-myometrial junctions [17]. A similar approach has also identified myometrial stem cells at the periphery of the longitudinal muscle layer [18]. The myometrium in the mouse uterus harbors cells with a quality of stemness [18]. Myocyte proliferation increases in the uteri of pregnant mice, and proliferated myocytes exhibit a synthetic phenotype after parturition, becoming hypertrophic [15]. Myometrial hypertrophy only occurs in gravid uterine horns under hormonal influence [15].
An interesting observation in this study is that mesenchymal cell replenishment occurred from mesometrial autophagy-intact cells to anti-mesometrial autophagy-deficient cells. The remaining question is whether the direction of the cell movement is always the same. This question arises because our mouse model had a clear functional deficit on the anti-mesometrial side. Thus, such deficits may provide guidance for cell mixing and movement. Under normal conditions, when all the mesenchymal cells are functionally equal, cell mixing during tissue repair may follow a distinct pattern. Whether cell mixing has a specific pattern during uterine involution requires further investigation.