Introduction
The fallopian tube (FT), also known as the uterine tube, is an important part of the female reproductive tract. The FT plays a critical function in human reproduction by providing a specific cellular and molecular microenvironment to support gamete transport, fertilization, and early embryonic development [1]. Therefore, FT obstruction or epithelial dysfunction may result in fertility impairment [2,3]. Interestingly, human FT epithelial cells are recognized as part of the innate immune system, which could mediate inflammatory responses to potential invaders [4]. Studies have investigated Toll-like receptors (TLRs) as key players in inflammation and found that they are highly expressed by FT epithelial cells [5-7]. TLRs play an essential role in immune activation through the recognition of pathogen-associated molecular patterns (PAMPs) and endogenous damage-associated molecular patterns (DAMPs) [5,6,8]. The interaction of sperm with the FT may affect sperm storage, motility, survival, and capacitation [9]. Sperm interaction with FT epithelial cells is a complex process that could induce biochemical and physiological alterations. Moreover, this process leads to the activation of innate immune mechanisms, such as TLR signaling [5,10]. In general, TLRs play a major role in the immune interaction between FT and sperm [11], sperm capacitation, fertilization, and pregnancy [6,11].
Despite advances in technology in the field of infertility treatment, the implantation success rate in embryo transfer cycles ranges from 26% to 45% [12]. Repeated implantation failure (RIF) is defined as implantation failure after the transfer of at least four high-quality embryos in at least three cycles. Since sperm provides half of the genetic material of the embryo, studies have investigated its role in RIF patients [13]. Sperm DNA integrity is an influential factor in embryo development and reproductive outcomes [14,15]. Although the sperm DNA fragmentation index (DFI) may be associated with an increased risk of miscarriage, its relationship with RIF has not been proven, and this issue needs further investigation [16]. Evidence has demonstrated that the sperm DFI is significantly elevated in infertile couples with unexplained or idiopathic infertility [17-19]. In this context, it has been shown that the DNA integrity of spermatozoa that attach to FT cells is higher than spermatozoa that do not attach [20]. Conspicuously, FT epithelial cells are actively able to distinguish normal and abnormal sperm chromatin [21]. Hence, this study aimed to evaluate the effects of sperm with high sperm DNA fragmentation on the expression of TLRs, adaptor molecules, and cytokines in FT epithelial cells through the polymerase chain reaction (PCR) array method.
Methods
1. Patient selection
This study was approved by the Institutional Review Board of the Ethics Committee of Royan Institute (Reference number: IR.ACECR.ROYAN.REC.EC/91/1084). Written informed consent was obtained from all patients.
Ten men with a history of unexplained RIF, a normal standard semen analysis (according to the 2010 World Health Organization [WHO] criteria) and a DFI >30% comprised the high-DFI group, and 10 normozoospermic men with at least one child and DFI <30% comprised the control group. Patients with a medical history of varicocele, testicular atrophy, undescended testis, genitourinary infection, chronic prostatitis, cigarette smoking, and alcohol consumption were excluded from the study.
2. Cell line (OE-E6/E7) and sperm preparation
The OE-E6/E7 cell line was cultured in Dulbecco’s Modified Eagle’s Medium/F12 (Invitrogen, Paisley, UK) with 1% penicillin and streptomycin (Sigma-Aldrich, Poole, UK), L-glutamine (Invitrogen, Waltham, MA, USA) and 10% fetal bovine serum (Invitrogen, Waltham, MA, USA). The cells were incubated in a 37°C incubator in an atmosphere of 5% CO2 in the air. Semen samples were analyzed according to the WHO guidelines and sperm preparation was performed by density gradient centrifugation and the swim-up method.
3. Sperm chromatin structure assay
The sperm chromatin structure assay was used to determine the DFI value. An aliquot of unprocessed semen containing 1-4 million sperm/mL was diluted with Tris-HCl, NaCl, and ethylenediaminetetraacetic acid (EDTA) (TNE) buffer. The cell suspension was treated with an acidic detergent solution (pH 1.2) comprising 0.1% Triton X-100, 0.15 mol/L NaCl, and 0.08 mol/L HCl for 30 seconds. The cells were then stained with 6 mg/L purified acridine orange (AO) (Polysciences, Warrington, PA, USA) in a phosphate citrate buffer (pH 6.0). Then, chromatin damage was determined by measuring the metachromatic change of AO fluorescence from green (associated with double-stranded DNA) to red (associated with single-stranded and denatured DNA). All analysis were performed by a technical expert [22] and a DFI >30% was considered high [23,24].
4. Sperm co-culture with OE-E6/E7
First, OE-E6/E7 cell culture was performed in 6-well culture plates until confluency. After sperm preparation in both groups (high-DFI and control), 1×103 sperm in each group were added to cultured cells separately [11]. The OE-E6/E7 cells were incubated with sperm for 24 hours. Then, the culture medium was collected. The OE-E6/E7 cells were washed with calcium and magnesium-free phosphate-buffered saline (PBS) and then were harvested with trypsin/EDTA (Invitrogen, Paisley, UK). The harvested cells were centrifuged at 300 ×g for 5 minutes and the pellets were used for RNA extraction.
5. RNA extraction and cDNA synthesis
RNA extraction was performed using an RNeasy mini kit (Cat. No: 73,304, Qiagen, Hilden, Germany), according to the manufacturer’s instructions. The quality and quantity of the extracted RNA were evaluated using a Nanodrop spectrophotometer device. cDNA synthesis was carried out using an RT2 first strand kit (Cat. No: 330,40, Qiagen) [25].
6. PCR array
The expression levels of 84 genes involved in the human innate immune response were evaluated by an RT2 profiler PCR array kit (Cat. No: PAHS-052A, Qiagen) and an RT2 SYBR Green ROX qPCR master mix (Cat. No: 330,502, Qiagen). Glyceraldehyde-3-phosphate dehydrogenase was considered as an endogenous control. The experiment was performed with a StepOnePlus Real-Time PCR System (Thermo Fisher, Waltham, MA, USA).
7. Statistical analysis
Data were expressed as mean±standard deviation. The normality of the data distribution was examined by the Kolmogorov-Smirnov test. The Mann-Whitney U test and the independent samples t-test were used for non-parametric and parametric analysis, respectively. All data were analyzed using SPSS ver. 22 software (IBM Co., Armonk, NY, USA), and a p<0.05 was considered as the significance level.
Results
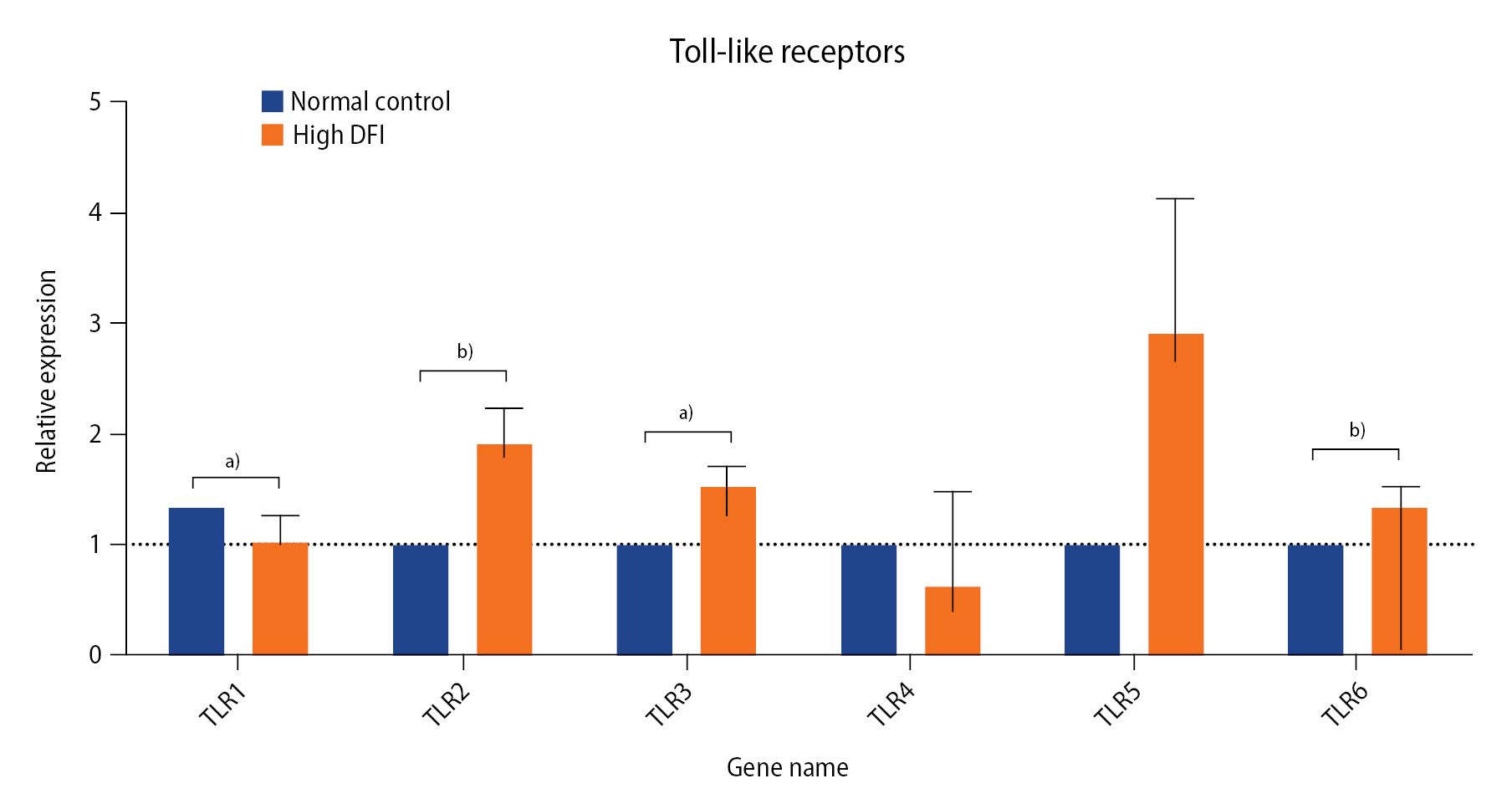
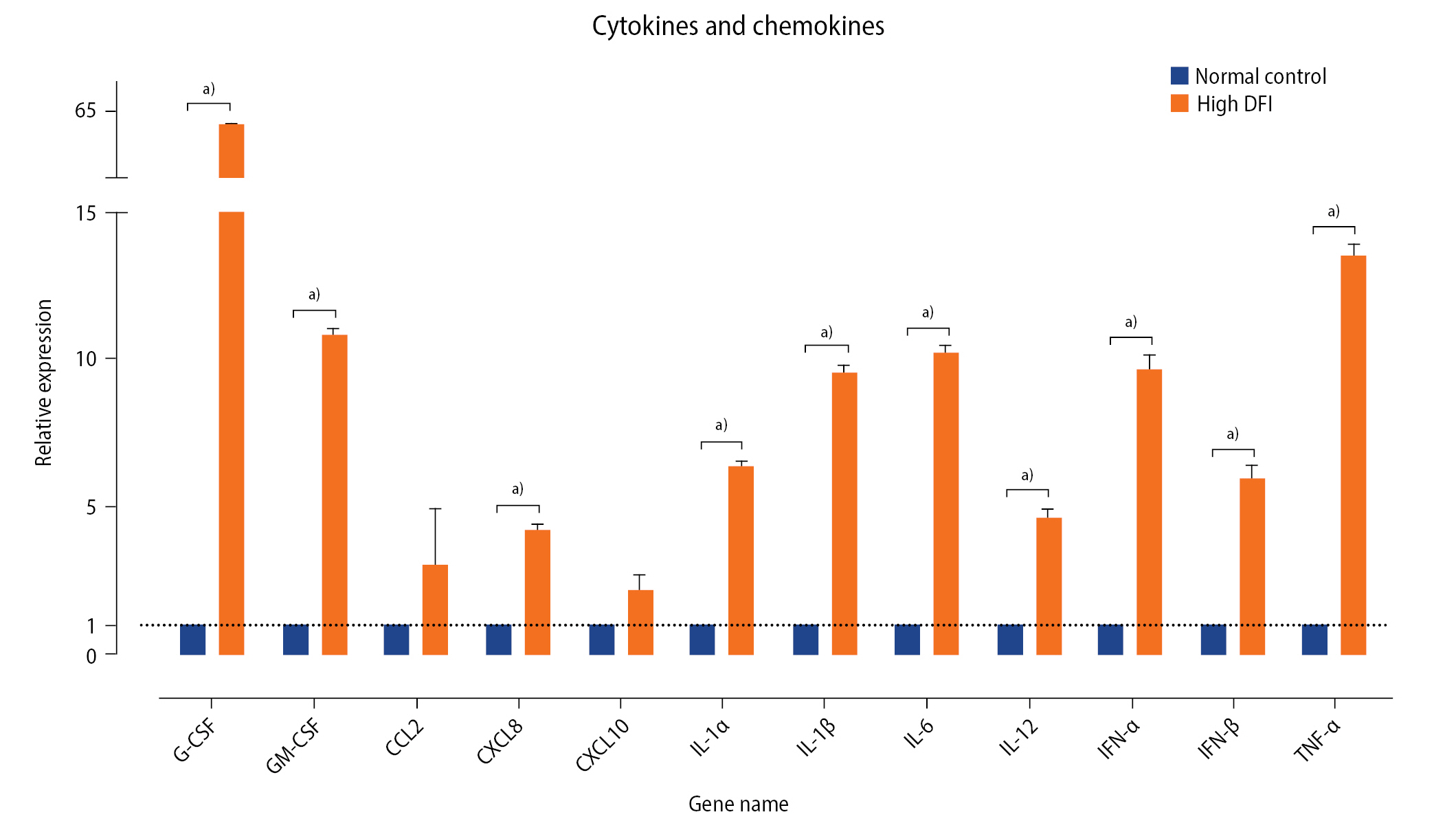
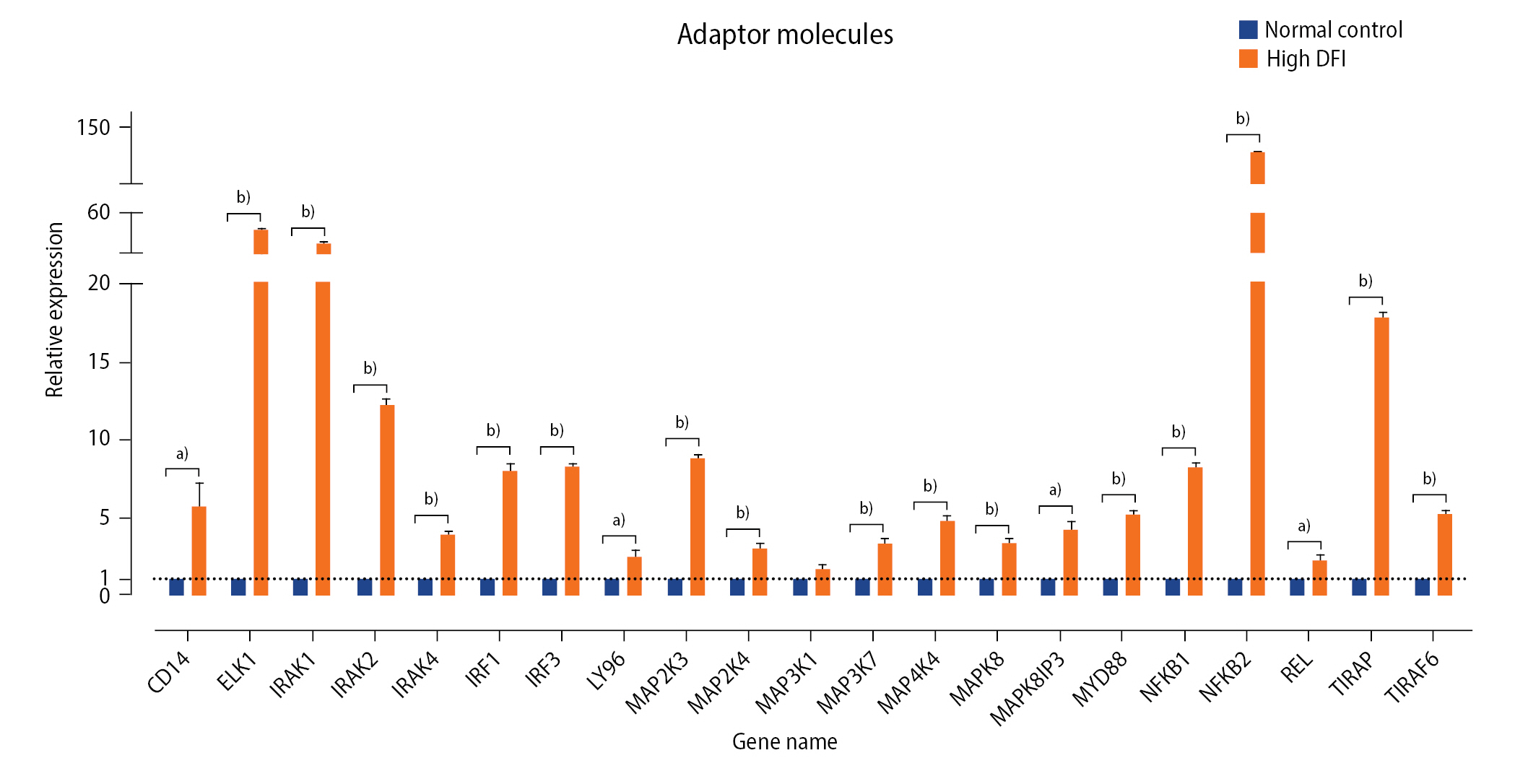
After 24 hours of co-culture, up to 90% of sperm were viable and unattached to the OE-E6/EE7 cells. Therefore, sperm were easily isolated from the cell line using PBS washes. A profiler PCR array was used to evaluate alterations in the expression of genes related to the innate and adaptive immune responses in the OE-E6/E7 cell line incubated with sperm from the two study groups (high-DFI and control groups). The genes with significantly upregulated expression in the high-DFI group in comparison with the low-DFI (control) group are shown in Figures 1-3 (p<0.05).
The expression levels of TLR-1, TLR-2, TLR-3, and TLR-6 in the high-DFI group were significantly higher than the control group (p<0.05) (Figure 1). The relative expression levels of proinflammatory cytokine and chemokine receptors, including interleukin 1α (IL-1α), IL-1β, IL-6, IL-12, interferon α (IFN-α), IFN-β, tumor necrosis factor α (TNF-α), C-X-C motif chemokine ligand 8 (CXCL8), granulocyte colony-stimulating factor (G-CSF), and granulocyte-macrophage (GM)-CSF were higher in the high-DFI group than in the control group (p<0.05) (Figure 2). Furthermore, the expression levels of several adaptor molecules, including cluster of differentiation 14 (CD14), ETS transcription factor ELK1 (ELK1), IL-1 receptor-associated kinase (IRAK1), IRAK2, IRAK4, interferon regulatory factor 1 (IRF1), IRF3, lymphocyte antigen 96 (LY96), mitogen-activated protein kinase kinase 3 (MAP2K3), MAP2K4, MAP3K7, MAP4K4, mitogen-activated protein kinase 8 (MAPK8), MAPK8IP3, myeloid differentiation primary response 88 (MYD88), nuclear factor kappa B subunit 1 (NFKB1), NFKB2, REL, Toll/interleukin-1 receptor domain-containing adapter protein (TIRAP), and TNF receptor-associated factor 6 (TRAF6), were significantly greater in the high-DFI group than in the control group (p<0.05) (Figure 3).
Discussion
The interaction of sperm and the FT is considered an essential event in the fertilization process, and it can activate an immune response. In this study, the expression of TLRs, adaptor molecules, and inflammatory cytokines and chemokines was evaluated in response to the interaction of sperm with high-DFI from RIF patients with a human FT epithelial cell line. The results showed that the interaction of high-DFI sperm from these patients with FT cells stimulated the innate immune system and specific inflammatory pathways.
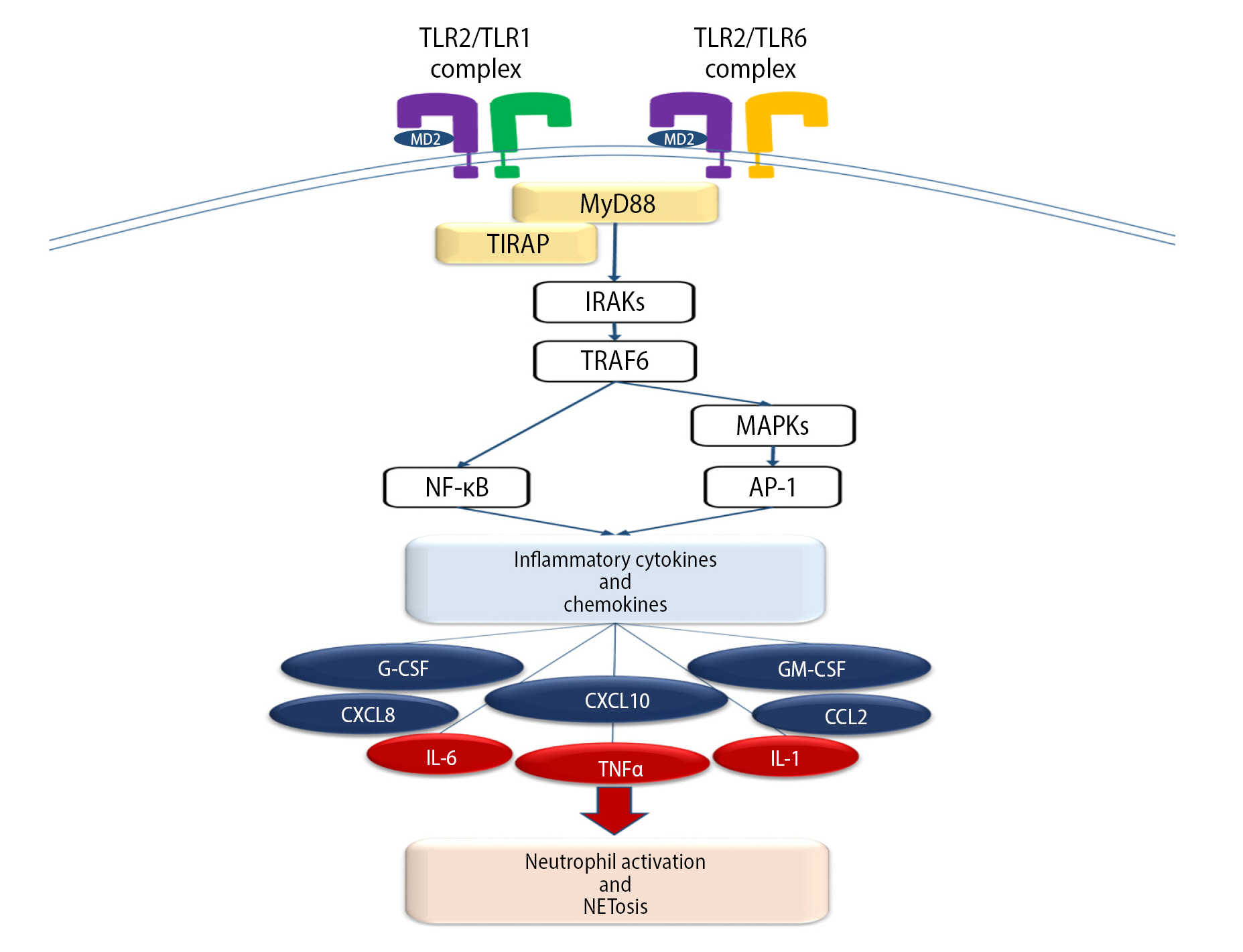
The innate immune system, as the first line of defense, plays an influential role by recognizing PAMPs and DAMPs through TLRs in FT epithelial cells [26]. Several immune pathways are potentially involved in the interaction of high-DFI sperm with the FT. This study revealed that the expression of TLR-1, TLR-2, and TLR-6 were significantly higher in the high-DFI group than in the low-DFI group (Figure 1). Existing evidence indicates that TLR-2 can form a heterodimer with TLR-1 or TLR-6. These complexes can recognize DAMPs, which are released from damaged cells. Following TLR signaling, two main pathways—the MyD88 and TIR-domain-containing adapter-inducing interferon-β (TRIF)—are activated. In particular, TLR-1, TLR-2, and TLR-6 signaling is mediated by the MyD88-dependent pathway [11]. The intracellular part of these complexes binds to cytosolic adaptor molecules, including MyD88 and TIRAP. Finally, signal transduction occurs through IRAK1. Subsequently, the TLR signaling pathway leads to the activation of activating protein-1 (Ap-1) and nuclear factor kappa light chain enhancer of activated B cell (NF-κB) transcription factors, which regulate the production of inflammatory cytokines and chemokines, including IL-1, IL-6 and, TNF-α, G-CSF, GM-CSF, CXCL10, CXCL8 (IL-8), and C-C motif chemokine ligand 2 (CCL2) (Figure 4) [27,28].
Our study results demonstrated that all transcription factors involved in the MyD88-dependent pathway were significantly increased in the high-DFI group, which may indicate the effect of high-DFI in stimulating this inflammatory pathway (Figures 2 and 3). Although these factors play an important role in fertility, their upregulated expression makes the FT conditions unfavorable for successful fertilization and healthy pregnancy. For instance, IL-6 plays a vital role in sperm capacitation and embryo implantation; however, the upregulation of IL-6 can reduce the ciliary activity of the FT epithelium and lead to defective embryo implantation in the FT [29]. IL-1 also causes destruction of the cilia in pathological conditions [30]. According to the evidence, a high concentration of IL-1 leads to the impairment of fertilization in a sperm penetration assay and zona pellucida assay and inhibits mouse embryo development [31]. Furthermore, the upregulation of TNF-α could reduce sperm motility and prevent sperm/mucus interaction [32].
Many studies have been conducted on the relationship between male infertility and RIF, but a connection has not been securely established. It is possible that high-DFI in sperm may lead to defects in embryo implantation through a negative effect on the FT. In a previous study, we cultured semen samples of 10 healthy men with the OE-E6/E7 cell line and examined the expression of 84 cytokines in the cell line using quantitative PCR. That study showed that sperm exhibited changing expression levels of cytokines, chemokines, and growth factors [33]. Furthermore, in another study, we cultured the sperm samples of 10 men with unexplained infertility and high-DFI compared to semen samples of 10 healthy donor men with OE-E6/E7 cell lines, and quantitative PCR analysis showed that DNA damage-induced activation of the TLR signaling pathway in human FT led to the upregulation of inflammatory cytokines and chemokines [11].
Oxidative stress, apoptosis, and chromatin remodeling are three major causes of sperm DNA fragmentation. In patients with high sperm DNA fragmentation, DAMPs are released from damaged spermatozoa, which can activate the immune response [34]. The FT epithelial cells may respond to these molecules through TLRs, leading to inflammatory cytokine secretion, which may orchestrate the infiltration of granulocytes, especially neutrophils [27,28].
As Figure 2 shows, IL-1 and TNF-α expression was increased in the high-DFI group; these molecules promote the expression of CXCL8 by endothelial cells [35]. However, CXCL8, also known as IL-8, regulates immune cell migration, especially the infiltration of neutrophils [27,36]. Neutrophils, the major granulocytes in the FT, respond to foreign molecular patterns through signaling pathways, antimicrobial agents, cytotoxic components, and proinflammatory cytokine production [37]. Interestingly, neutrophils can be found in the lumen of the FT through transepithelial migration. In general, neutrophils are capable of producing high amounts of reactive oxygen species, which exert a negative effect on sperm motility and fertility [38]. The activation of neutrophils can also lead to NETosis and apoptosis (Figure 5). NETosis is considered as a cell death model that is different from necrosis and apoptosis. Neutrophil extracellular traps (NETs) are structures consisting of decondensed chromatin and expanded nuclear material that combine with cytoplasmic components after the nuclear membrane decomposes. They are then released following the rupture of the plasma membrane, leading to the trapping and killing of microorganisms [39-41]. It has been shown that the incubation of peripheral neutrophils with equine sperm resulted in severe NET formation. It also has been observed that the repeated deposition of sperm in the presence of neutrophils could reduce fertility potential [42]. These plausible immuno-inflammatory mechanisms may be viewed as a potential factor in male infertility. Since the pathophysiological identification of patients with RIF can guide the treatment of patients and increase the success rate, more research on the effects of sperm with high-DNA fragmentation on the female reproductive system and a better understanding of the immune-inflammatory mechanism and neutrophil function in these patients are needed.
In conclusion, this study provides novel insights into the role of the innate immune responses and TLR signaling pathways in men with history of unexplained RIF. The MyD88-dependent pathways of TLR-1, TLR-2, and TLR-6, as well as neutrophil activation, are inflammatory pathways activated by high-DFI sperm that could be considered as a potential immunopathogenesis of male infertility.