The oocyte pool is the primary determinant of reproductive lifespan
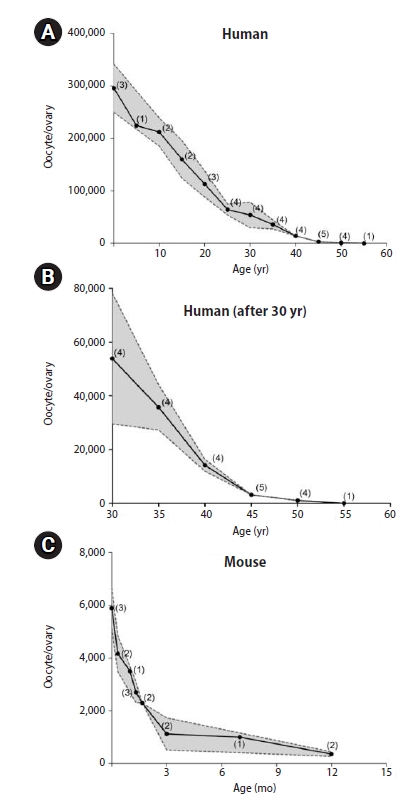
In women of reproductive age, the size of the oocyte pool (the total number of oocytes in the ovaries) is the primary predictor of fertility [1]. It is well established that the oocyte pool can be as large as a few million during the fetal stage, but decreases significantly to a million or less at birth. Here, we report the predicted patterns of change in the oocyte pool in humans and mice. The human predictions were made based on collective study reports published since 1960 and those that are currently available via PubMed (Figure 1A) [2-8]. Similarly, predictions were also made in mice because they are widely used as animal models for researching human fertility (Figure 1C) [9-18]. Briefly, the predictions in humans indicate that an ovary contains approximately 300,000 oocytes at birth (600,000 per person). The oocyte pool decreases in size throughout the entire reproductive lifespan, down to half by the onset of puberty and one-third by the age of 20. Interestingly, substantial variation in the oocyte pool size is seen by the age of 40 (Figure 1B), but this variation subsequently disappears. Eventually, the human oocyte pool becomes close to zero by the age of 50 years, when most women reach menopause. A similar declining pattern of the oocyte pool is also seen in mice, indicating that there is a regulatory mechanism that is common to both species.
There is a continuing trend worldwide for the current generation of women to plan to bear children at a considerably later time of life than in previous generations. This trend creates a risk that when the time to have a child arrives, the oocyte pool may not be large enough to support fertility even with help from assisted reproductive technology. No method currently exists for precisely assessing the oocyte pool. Instead, a few methods are used as clinical assessments of the oocyte pool, including measurements of blood levels of anti-Müllerian hormone (AMH) alone or together with follicle-stimulating hormone, the levels of which are affected by the number of follicles in the ovary and the amount of estradiol synthesized by the follicles, respectively [19,20]. AMH is a peptide hormone produced by the granulosa cells of the primary and secondary follicles and released into the bloodstream. Therefore, the blood level of AMH is used as a proxy of the size of the oocyte pool, and this method has been adopted by most in vitro fertilization (IVF) clinics to assess the size of their patients’ oocyte pool. Low or high blood AMH levels are used as a clinical marker to predict whether ovulation can be induced in an IVF patient or to determine how many oocytes can be retrieved upon inducing ovulation. However, neither AMH nor follicle-stimulating hormone levels are reflective of the actual size of the oocyte pool. Indeed, these levels do not change for the entire fertile lifespan even though the oocyte pool continues to decrease in size; they only begin to change when the oocyte pool becomes so small that not enough primordial follicles can be recruited to the growth phases. Interestingly, in a survey performed with 796 infertility clinics asking which test or factor best predicts fertility, 80% of respondents stated that age was the best predictor [21]. Why age? The oocyte pool decreases nonstop even when a woman’s fertility is at its peak or she is in her teens or 20s. As of 2021, no consensus exists regarding the cause(s) of the continued decline of the oocyte pool, but it is believed to be a multifactorial phenomenon. In the following sections, we summarize reported causes of the decline that are backed by experimental or observational data. Additionally, the hypothesis that ovulatory inflammation is a contributing factor to the decline of the oocyte pool will be discussed.
Contributing factors to the decline of the oocyte pool
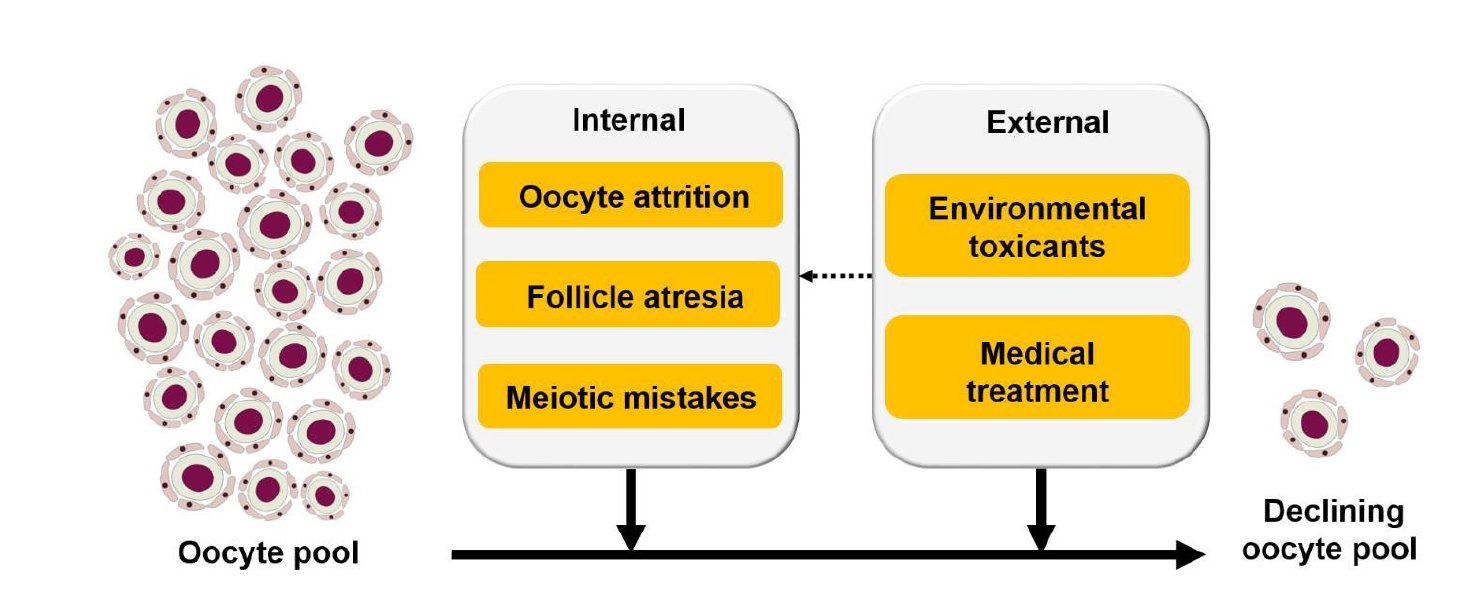
The peak size of the oocyte pool is established during fetal development, specifically during the transitional period of mid-to-late gestation [22]. Upon reaching the peak, the pool begins to decrease, primarily via a process called oocyte attrition, throughout the rest of the gestational period [23,24]. After birth, the oocyte pool continues to decrease due to a variety of factors including, but not limited to, follicular atresia, meiotic errors, ovulation, environmental toxicants, medical drugs, and possibly some other factors that are yet to be discovered (Figure 2).
1. Perinatal oocyte loss
Immediately prior to or after birth, the ovary loses oocytes via oocyte attrition, which is mediated by immune cells infiltrating the fetal ovaries [25] or apoptosis triggered by DNA breaks [26,27]. Furthermore, autophagy, ferroptosis, and necrosis are also known to cause oocyte demise. Autophagy is a cell recycling pathway that involves the fusion of organelles with lysosomes for degradation [28-30]. Ferroptosis is a type of programmed cell death involving intracellular iron and oxidative phospholipids [22,31]. Necrosis is an inflammatory cell death process that results in swelling and eventually cell rupture [32].
2. Follicular atresia
After birth, a subset of dormant primordial follicles is recruited into a growth phase, which continues throughout the entire reproductive lifespan. Some of them grow and develop into preantral follicles, and eventually release oocytes via ovulation. However, the majority of the recruited follicles die during the growth phase; they eventually degenerate and are removed from the ovary. This phenomenon is called follicular atresia and is often triggered by the death of a granulosa cell in a follicle [22], eventually leading to the death of most granulosa cells and oocytes. Decreased estrogen production in the follicle or a milieu with increased androgen levels is a known cause of the initial death of granulosa cells [33-36]. In addition, erroneous signaling between the oocyte and granulosa cells also causes mis-regulation in the meiotic arrest of oocytes and follicular atresia [37-40].
3. Environmental toxicants
Humans are constantly exposed to environmental toxicants via inhalation, ingestion, or contact. Recent studies have shown that an alarming number of environmental toxicants can directly or indirectly damage oocytes. For example, oocytes in an ovary that is exposed to cadmium die due to an increased level of reactive oxygen species (ROS), subsequent DNA damage, and mitochondrial disruption in the oocytes [41]. Di(n-butyl) phthalate, an industrial plasticizer and stabilizer, kills oocytes by activating apoptosis pathways upon exposure in animals [42]. Nonylphenol, a commonly used industrial material similar to laundry detergents, has been shown to alter the expression of more than 800 genes in mouse oocytes, increase ROS levels, and eventually eliminate oocytes [43]. Further, a variety of endocrine-disrupting chemicals bind to steroid receptors (e.g., estrogen or progesterone receptors), synergistically disrupt hormonal balance, and directly or indirectly harm oocytes [44-46]. Unlike oocyte attrition and follicular atresia, which are mostly triggered by internal factors, environmental toxicants are from external sources and therefore preventable to some degree.
4. Medical treatments
Multiple drugs are used to cull harmful cells, particularly in cancer patients. These drugs primarily target cell-cycle regulators, making the ovary a vulnerable organ to such medical treatment. When given to a patient, these drugs inevitably impact cells in the ovary because a substantial proportion of ovarian cells undergo cell proliferation (granulosa cells in follicles at various stages) and meiosis (oocytes). Indeed, it is well known that young female cancer patients who receive chemotherapy suffer from premature ovarian insufficiency (POI) [47,48]. Chemotherapeutic drugs such as cyclophosphamide, cisplatin, vincristine, and doxorubicin are all reported to induce POI [49-51]. In addition to targeting cell cycle regulators, some chemotherapeutic drugs “burn out” the oocyte pool. For example, cyclophosphamide stimulates follicle activation in oocytes and granulosa cells, thereby pushing an excessive number of primordial follicles into a growth phase and decreasing the oocyte pool [47]. Doxorubicin triggers the DNA-damaging TAp63α-Cleaved-CASP3 pathway to induce apoptosis in the oocytes of primordial follicles [48]. Radiation therapy is also toxic to oocytes. It increases ROS production in patients, which induces DNA damage either directly or via lipid peroxidation or by increasing the expression of a pro-apoptotic protein, p53 upregulated modulator of apoptosis, in oocytes or granulosa cells [52]. Cryopreservation of oocytes or ovarian tissues has been used to persevere oocytes for later use before chemotherapy [50]. Further, antioxidants such as melatonin are now used to quench ROS before radiation therapy [53].
5. Meiotic errors
Meiotic errors can be caused by environmental toxicants or medical treatments. However, they can also happen by chance during normal oogenesis. Typical meiotic mistakes include failure to repair DNA breaks and defective synapsis [26], abnormal spindle assembly [54], and further chromosome nondisjunction [55]. Surprisingly, 20%–25% of human oocytes obtained in IVF clinics showed aneuploidy [56], and the ratio increases as women become older [57]. To control the quality of the overall oocyte pool, certain meiotic checkpoint mechanisms sacrifice abnormal oocytes and decrease the total oocyte number [54,57]. However, these checkpoints are less stringent in females than in males [57]. Thus, some oocytes with meiotic errors can still be fertilized, leading to chromosome-related diseases/syndromes such as Down syndrome in newborns [58,59]. Nonetheless, oocytes can find a way to efficiently repair DNA double-strand breaks and protect the genetic integrity of offspring if apoptosis is inhibited [60,61].
Ovulatory inflammation: a contributing factor to the decline of the oocyte pool?
Ovulation is the process of releasing oocyte(s) from the ovary. It is triggered by a preovulatory surge of luteinizing hormone (LH) and recurs every reproductive cycle [62]. In the entire reproductive life span, a woman releases only 300–400 oocytes [63], a negligible subset of oocytes compared to the number of oocytes a female is born with. In the previous section, a few well-established causes of oocyte losses were reviewed. In this section, we will discuss another cause that may trigger oocyte loss: the acute inflammation that the ovary experiences at the time of ovulation. When a tissue undergoes acute inflammation, leukocytes quickly infiltrate the tissue and display a series of defensive behaviors with the aim of destroying foreign invaders, such as infectious microorganisms or harmful substances. However, these defensive actions inevitably damage the tissue itself. Overwhelming evidence indicates that the ovary experiences acute inflammation when it undergoes a cascade of ovulatory processes, suggesting that a similar tissue-damaging event may happen in the ovary at the time of ovulation. Therefore, the ovarian cells, including primordial germ cells, may be damaged and eventually removed from the ovary at each ovulation, accounting for a significant portion of oocyte loss during the reproductive lifespan. Herein, we will discuss this possibility.
1. Ovulation is accompanied by acute inflammation
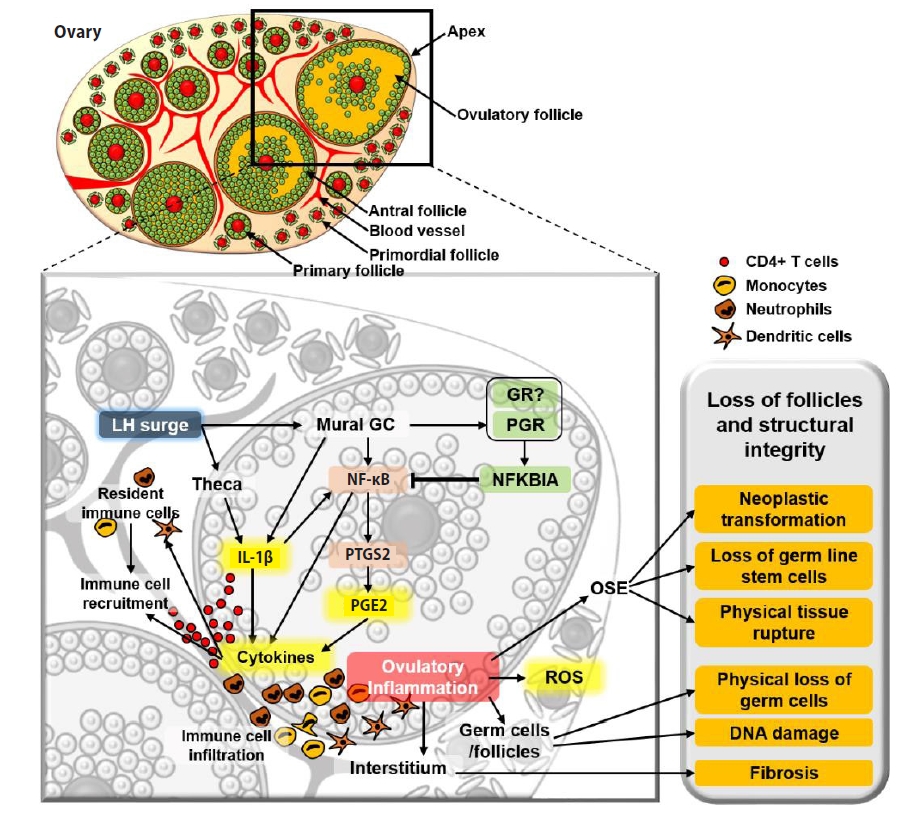
The idea that ovulation is an inflammatory process was originally introduced in the early 1980s by Espey [64], who presented a striking similarity between a general inflammatory process and the events that take place at the time of ovulation. Since then, this idea has gained popularity through extensive observational and experimental data that were recently reviewed by Duffy et al. [65]. The most obvious evidence that the ovary undergoes acute inflammation is that the number and density of proinflammatory leukocytes increase in the ovary immediately after the LH surge [66-69], like any other tissue that undergoes acute inflammation. The LH surge-induced leukocyte infiltration is mediated by pro-inflammatory cytokines and ROS that are either released by ovarian cells or resident leukocytes [64,70-74]. For example, one of the pro-inflammatory cytokines, interleukin 1 beta (IL-1β), is expressed and secreted from theca cells, granulosa cells, and leukocytes [75,76]. IL-1β subsequently induces the expression of prostaglandin synthase 2 (PTGS2) in granulosa cells [76]. PTGS2 is a rate-limiting enzyme in the synthetic pathway of prostaglandin E2 (PGE2) [77,78], a pro-inflammatory agent that impacts angiogenesis, blood flow, immune cell function, and tissue remodeling in the ovary [65]. Importantly, locally produced PGE2 stimulates the recruitment of a variety of proinflammatory cells to the ovary in a short time [79], as it does in other inflammatory sites [80], creating acute inflammation in the ovary.
2. Ovulatory inflammation and tissue damage
Acute inflammation causes oxidative stress and DNA damage in the resident cells, and this occurs in the ovary when it undergoes ovulatory processes [81-84]. As the ovary repeats ovulation, the ovarian tissue, including oocytes, may also be damaged by proinflammatory cells via engulfment and the proteases that the immune cells release [85]. Importantly, inflammatory cells and their secretions are primarily localized and execute their inflammatory actions around the blood vessels in the ovulatory follicles and medullary region. However, primordial follicles are sequestered in the cortex, the least vascularized region in the ovary. Therefore, presumably, the primordial follicles localized close to medulla, but not those far away from the inflammatory sites and in the cortex, may be damaged and lost at the time of ovulation (Figure 3).
In support of the suggested role of inflammatory cells in decreasing the oocyte pool, recent studies have pointed out that a pathological level of ovarian inflammation induces POI, which is caused by an accelerated decline of the oocyte pool [86-89]. For instance, women with chronic systemic inflammation caused by an autoimmune disease (e.g., Addison disease) or by pathologically elevated production of inflammatory cytokines have a significantly higher likelihood of developing POI [87,90-93].
In addition to its direct impact on oocytes or follicles, inflammation may indirectly accelerate the decline of the oocyte pool by inducing fibrosis (deposition of connective tissue), which is generally observed in aged ovaries [94,95]. Indeed, fibrosis is a part of the natural aging process in the ovary and likely contributes to the decline of the oocyte pool, as fibrous tissues may interfere with normal ovarian dynamics, as well as impair follicular survival, recruitment, and growth (Figure 3).
3. Polycystic ovarian syndrome, contraception, and the oocyte pool
If it is true that ovulatory inflammation accounts for the loss of a significant portion of oocytes, and therefore is a contributing factor to the decline of oocyte pool, one can predict that the oocyte pool decreases more slowly and therefore the reproductive lifespan would be longer in women whose ovulation frequency or number is less than that of average women. Women with polycystic ovarian syndrome (PCOS) do not ovulate at all or have a much lower ovulation frequency than those without the syndrome. As a result, women with PCOS experience a significantly lower level of ovulatory inflammation. Indeed, women with PCOS have a larger ovarian pool and exhibit a slower rate of follicular atresia than infertile eumenorrheic women [96-98]. Another example of less ovulation can be seen in women who take contraceptive pills. Unfortunately, there is no direct evidence that taking contraceptive pills may delay the oocyte-pool decline. However, it is well established that women who take contraceptive pills have a lower probability of developing ovarian cancer than those who do not [99-101], likely because the ovarian tissues have less exposure to ovulatory inflammation.
In summary, the literature clearly shows that the causes of the continuous decline of the oocyte pool throughout the entire reproductive lifespan are multifactorial. Some factors are intrinsic to the ovary (oocyte attrition, meiotic errors, follicular atresia) and unavoidable, while others are external and preventable (environmental toxicants, drugs). In addition, here, we discuss an additional potential intrinsic factor: ovulatory inflammation. The question remains of why females are born with excess oocytes. Might it be to ensure that they maintain fertility for a sufficient period when the ovary continues to lose oocytes due to the aforementioned reasons? Males are born without sperm in their gonads. Instead, their gonads contain germ cell stem cells, from which sperms are continuously produced for a few decades in humans, with the stem cells serving a reservoir for continued production of the male germ cells. The female gonads, however, do not contain germ cell stem cells; instead, ovarian egg production has to rely on the finite number of oocytes with which they are born. Therefore, having an excess number of oocytes at the time of birth may be a mechanism to ensure egg production for an extended period and long-term fertility in humans.