Introduction
The term in vitro maturation (IVM) refers to the maturation of immature oocytes retrieved from antral follicles with minimal or no gonadotropin stimulation [1]. Developed as early as 1935 by Pincus and Enzmann, this technique—although it seemed promising—did not come to the limelight like conventional in vitro fertilization (IVF). IVM was initially done in natural cycles and with the advent of better protocols, the option of follicle-stimulating hormone (FSH) priming along with luteinizing hormone (LH)/human chorionic gonadotrophin (hCG) triggering has been advocated to enhance oocyte competence [2]. Irrespective of FSH/LH/hCG priming, the outcomes of IVM for oocytes with regard to fertilization/implantation rates have been bleak in comparison with conventional stimulation cycles [3]. The suboptimal embryo quality obtained from IVM oocytes could be due to asynchrony between nuclear and cytoplasmic maturation at the cellular level [4]. Nuclear maturation comprises re-initiation of meiosis in oocytes arrested at prophase I, which is heralded by the loss of the nucleolar membrane (germinal vesicle [GV] breakdown) and ends in the completion of meiosis I with a concomitant expulsion of the first polar body [5]. Cytoplasmic maturation encompasses all the events at the organelle level along with various epigenetic modifications that prepare oocytes for subsequent fertilization and embryo development [6]. Furthermore, cytoplasmic maturation cannot be quantified by events as nuclear maturation and is therefore poorly understood. One approach that has been postulated to enhance oocyte competence is to promote nuclear and cytoplasmic synchrony by delaying nuclear maturation while simultaneously allowing cytoplasmic maturation to manifest [7]. This might possibly lead to the development of good-quality embryos. In vivo, cyclic adenosine monophosphate (cAMP) is the main meiotic inhibitor that keeps the oocyte in an arrested state (prophase I) until the hours preceding ovulation. The cAMP generated in granulosa cells is also transported via gap junctions [8,9]. The maintenance of an elevated intra-oocyte cAMP level maintains meiotic arrest and keeps the oocyte in its dormant state. Synchrony can therefore be achieved by a prematuration culture that maintains higher levels of cAMP and inhibits the meiotic cycle within the oocyte in vivo [10,11]. Levels of cAMP are maintained by inhibiting the enzyme-phosphodiesterase (PDE) or by upregulating adenyl cyclase. Although nuclear and cytoplasmic maturation are stepwise processes, their coordination is essential for the development of competent oocytes. Various studies have shown that about 5% to 20% of oocytes retrieved in controlled ovarian stimulation (COS) cycles are immature [1,12]. Some of these oocytes have the potential for spontaneous maturation during in vitro culture, and are used for intracytoplasmic sperm injection (ICSI). However, immature oocytes are usually discarded due to the possibility of an increased rate of miscarriages or risk of anomalies [13,14]. In patients with a cohort of asynchronized follicles, where there is a higher proportion of immature oocytes, the use of these oocytes is essential to increase the number of injectable oocytes for ICSI and eventually to generate more embryos for transfer. Studies have explored various prematuration cultures of immature oocytes, probing the use of PDE-3 inhibitors (PDE-3Is) at an ideal concentration and duration of exposure [15]. The main challenge in developing IVM culture systems is to adopt the ideal culture conditions required in a particular stage of oocyte dependency [16]. PDE-3 is located in oocytes and PDE-4 in granulosa cells. Hence, the PDE-3I cilostamide is unique as it exerts its effects on oocytes specifically and has no effect on granulosa cells [17,18]. Hence, it causes a delay in nuclear maturation without hindering cytoplasmic maturation [19]. The objective of this study was to assess the developmental potential of GV oocytes aspirated in COS cycles that were subjected to IVM after prematuration culture with cilostamide. We also attempted to study the impact of cilostamide exposure at the GV stage on the morphology of meiosis II (MII) oocytes and subsequent embryo quality.
Methods
1. Ethical approval
The study was conducted at a tertiary care reproductive medicine unit in India. It was approved by the Institutional Review Board and the Nitte Central Ethics Committee (NU/CEC/2018/0191 NU/CEC/2020/2069). Informed consent was obtained from patients prior to the laboratory access of oocytes and sperm.
2. Study population and oocyte source
In total, 994 oocytes were collected from 63 patients, of which 307 were GV oocytes. Thirty-seven morphologically abnormal GV oocytes were excluded from the study. The present study comprised 270 viable GV oocytes obtained from women aged 25–35 years, who underwent COS for IVF/ICSI at Milann Fertility Center from April 2018 to March 2020. Patients with a minimum of four GV oocytes were included in the study. Only one treatment cycle per patient was incorporated in the study.
3. Stimulation protocol
COS was started on day 2 or 3 of the menstrual cycle after a baseline transvaginal ultrasound scan (Voluson P6 with a 4-8 MHz vaginal probe) and hormonal assessment (estradiol [E2], progesterone, and LH if deemed necessary). Gonadotropins included either recombinant FSH, (Recagon, Organon, Oss, The Netherlands; Gonal F, Merck Serono, Modugno, Italy) or human menopausal gonadotropin (Menopur; Ferring Leciva, As, Czech Republic). The starting dose was calculated based on age, body mass index, and antral follicle count, and ranged from 112.5 to 187.5 IU daily for 4 days. The ovarian response to stimulation was monitored on the fifth day of stimulation with transvaginal ultrasound scan and serum E2, LH, and progesterone measurements, and the dose of gonadotropins was adjusted accordingly. A gonadotropin-releasing hormone antagonist (0.25 mg; Cetrotide, Merck, Darmstadt, Germany) was given daily subcutaneously based on a flexible antagonist protocol when the dominant follicle was ≥12–14 mm in diameter or the serum E2 level was >350–400 pg/mL until the day of triggering. Triggering was performed when at least 3 follicles reached ≥17 mm in diameter. Oocytes were aspirated transvaginally 35–36 hours post-triggering under intravenous sedation and ultrasound guidance. A single-lumen oocyte retrieval needle (Vitrolife, Vastra Frolunda, Sweden) was used.
4. Laboratory protocol
1) Preparation of cilostamide
The PDE-3I cilostamide (Cayman Chemical, Ann Arbor, MI, USA) was used for prematuration culture. A stock solution was prepared by dissolving cilostamide in crystalline form with dimethyl sulfoxide. A final concentration of 1 μM cilostamide was added to the prematuration culture with the intention of achieving efficient meiotic inhibition and maximum reversibility of inhibition.
2) Preparation of cumulus-oocyte complexes
Cumulus-oocyte complexes (COCs) were incubated in the culture medium (G-IVF-PLUS, Vitrolife) covered with mineral oil (Ovoil, Vitrolife) at 37°C in 6% CO2 for 2–3 hours. Cumulus cells were removed after exposure to G-Mops (3-N-morpholino-propane sulfonic acid) buffered medium containing 80 IU/mL hyaluronidase (Hyase, Vitrolife) for 30 seconds with the help of a glass Pasteur pipette (Humagen Fertility Diagnostics, Charlottesville, VA, USA). The nuclear status of the denuded oocytes was then assessed. Oocytes that were observed to have released the first polar body were considered mature and used for ICSI. Immature oocytes with a prominent nucleus (GVs) were assigned to the study and taken up for the prematuration culture.
5. Experimental setup
1) Prematuration culture
The denuded GV-stage oocytes of each patient were distributed into two groups (experimental and control group) and subjected to two different culture conditions. All immature oocytes were cultured individually in 25-μL drops of medium overlaid with oil in a humidified atmosphere at 37°C in 6% CO2. The oocytes in the experimental group were placed in an IVM medium (Oocyte Maturation Medium; Cooper Surgical/SAGE, Trumbull, CT, USA) supplemented with 5 mg/mL serum protein substitute and FSH/LH (75 mIU/mL for each; Serono, Mississauga, ON, Canada) supplemented with 1 μM cilostamide, while the oocytes in the control group were placed in an IVM medium without cilostamide. After culturing these GV-stage oocytes for 6 hours [20], the cilostamide was removed from the experimental group. These oocytes were then transferred to inhibitor-free IVM medium and cultured for 42 hours.
2) Maturation of oocytes
The oocytes were then classified as GV, MI, or MII. ICSI was performed on MII oocytes, and the GV oocytes were cultured up to 48 hours.
3) Semen samples for ICSI
Semen samples were collected at the time of oocyte retrieval. After liquefaction, a simple preparative method, such as the swim-up or double density gradient method, was used to prepare semen samples for the ICSI procedures [21,22]. Since the oocytes did not mature synchronously during culture, the semen samples were frozen in multiple vials and thawed accordingly for further insemination.
4) Intracytoplasmic sperm injection
Morphologically normal spermatozoa were injected into the GV-converted MII oocytes. Following ICSI, these two groups (experimental and control groups) of injected oocytes were then incubated in 25-μL drops of single-step medium (GTL; Vitrolife) for further development.
5) Fertilization assessment
Fertilization was assessed at 18 hours post-ICSI. Fertilization was determined by the appearance of two distinct pronuclei and two polar bodies.
6) Embryo culture system
The fertilized oocytes were cultured for 120 hours. Embryo development was observed at 24-hour intervals, which were continued until day 5 (120 hours). On days 3 and 5, the embryos were scored as per the Istanbul consensus [23]. Embryo grading was performed and validated by expert embryologists.
6. Statistical analysis
Data were analyzed using IBM SPSS ver. 23 (IBM Corp., Armonk, NY, USA). The differences between the treatment groups were analyzed using the chi-square test and when appropriate, the two-tailed Fisher exact test. A p-value <0.05 was considered to indicate statistical significance.
Results
The present study comprised 270 GV oocytes, of which 140 oocytes were selected for the experimental group and 130 oocytes for the control group (Figure 1). The quality and morphology of the GV oocytes were similar in both the experimental and control groups (p>0.05).
1. Blocking of nuclear maturation by prematuration culture with cilostamide
The GV oocytes in the experimental group were exposed to 1 μM cilostamide for 6 hours. The control group was free from cilostamide. After 6 hours of culture in cilostamide, it was observed that 92.1% of oocytes remained in the GV stage and only 6.4% of oocytes had converted to the MI stage. However, in the control group, 20.8% of GV oocytes converted to the MI stage. A statistically significant result was observed between the experimental and control groups (p<0.01) (Table 1). It was shown that cilostamide was effective at blocking nuclear maturation and promoting cytoplasmic growth. The number of MI oocytes showed a statistically significant difference between the experimental and control groups (p<0.001) (Table 1).
2. Culture system of GV oocytes after 6 hours: progression of meiosis at 26, 36, and 48 hours in oocytes subjected to prematuration culture with cilostamide (1 μM) followed by IVM culture
The GV oocytes were then cultured in IVM medium (without cilostamide) and were observed at 26, 36, and 48 hours (Table 2). The experimental and the control groups showed no significant differences at 36 hours and 48 hours, although a significant difference was found for GV oocytes at 26 hours (p=0.05). The experimental group showed the highest number of MII oocytes at 36 hours (25.8%), followed by 48 hours (13%) and 26 hours (10.8%). The control group also showed the maximum number of MII oocytes at 36 hours (21.5%), followed by 26 hours (14.6%) and 48 hours (7.9%). There was no statistically significant difference between the experimental and control groups (p>0.05). For the 57 oocytes that matured from 140 GV oocytes in the experimental group and the 48 oocytes that matured out of 130 GV oocytes in the control group, the maturation rate was 41% and 37%, respectively. The present study also observed an increased number of degenerated oocytes at 48 hours in the control group as compared to the experimental group (Table 2).
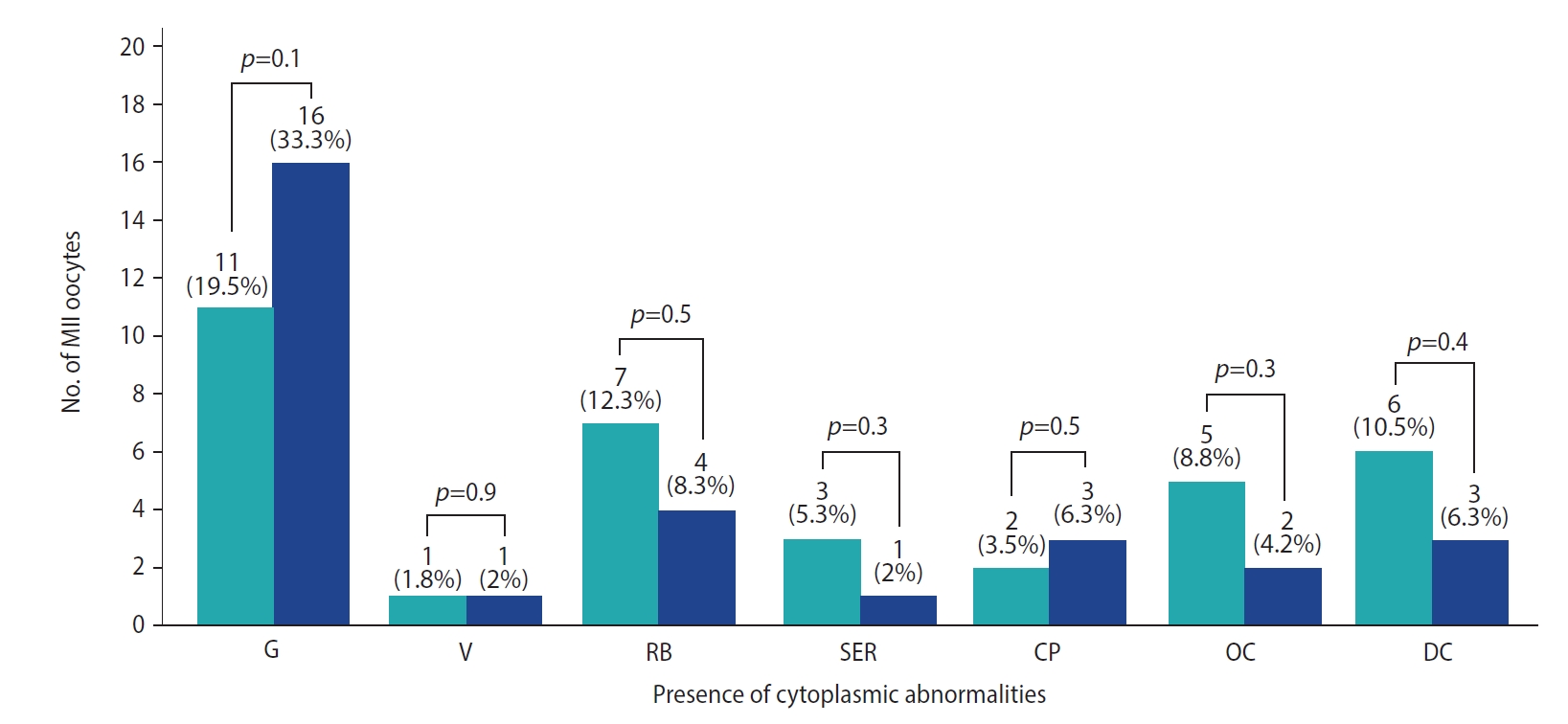
3. Morphological assessment of MII oocytes
Cytoplasmic abnormalities such as increased granulations, the presence of vacuoles, refractile bodies, smooth endoplasmic reticulum clusters, organelle clustering, and dark cytoplasm were found in the MII oocytes of both the experimental and control groups (Figure 2). However, the morphological features were similar in both groups (p>0.05). All the MII oocytes were then injected with morphologically normal sperm.
4. Fertilization of oocytes
The total number of MII oocytes obtained was 57 in the experimental group and 48 in the control group. Fertilization was confirmed with the appearance of two distinct pronuclei and two polar bodies in 35 oocytes (61.4%) in the experimental group and 22 oocytes (45.8%) in the control group. However, there were no statistically significant differences between the two groups (p>0.05) (Figure 1).
5. Cleavage of embryos
The fertilized oocytes in the experimental (n=35) and control (n=22) groups were further cultured. After 27 hours, the embryo culture showed a higher frequency of cleavage in the experimental group (88.6%) than in the control group (77.3%), although the results were statistically insignificant (p=0.2). Similar results were observed on day 2 and day 3 of embryo culture with regard to cleavage (p>0.05). However, optimal cleavage was more frequent in the experimental group (40%) than in the control group (27.3%), although this difference did not reach statistical significance (p=0.33) (Table 3).
Even though the cleavage rate was similar in both groups, the proportion of good-quality embryos (i.e., those with no apparent morphological abnormalities and few anucleate fragments) obtained was higher in the experimental group than in the control group.
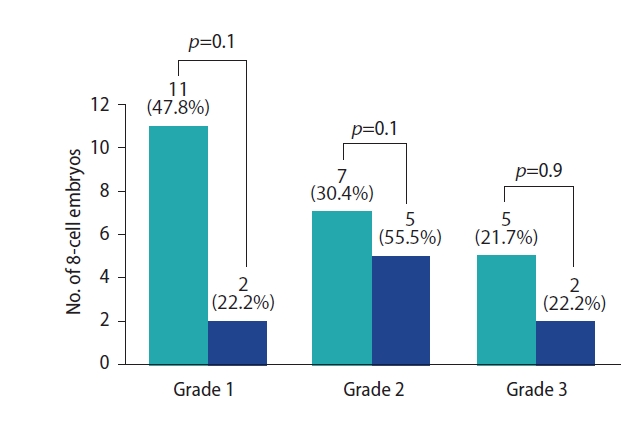
The experimental group showed the highest number of grade 1 embryos, followed by grade 2 and grade 3, whereas the control group showed the highest number of grade 2 embryos, followed by grade 1 and grade 3. However, the distribution of grades 1, 2, and 3 among the experimental and control groups was similar (p>0.05) (Figure 3).
6. Hourly cleavage status of embryos
The number of good-quality embryos was higher in the experimental group than in the control group, although this difference did not reach statistical significance. Considering the optimal time required to reach the cleavage stage, culturing for 26 hours and 36 hours resulted in the maximum number of embryos, whereas prolonged culture up to 48 hours did not give any advantage in terms of grade 1 embryos (Table 4).
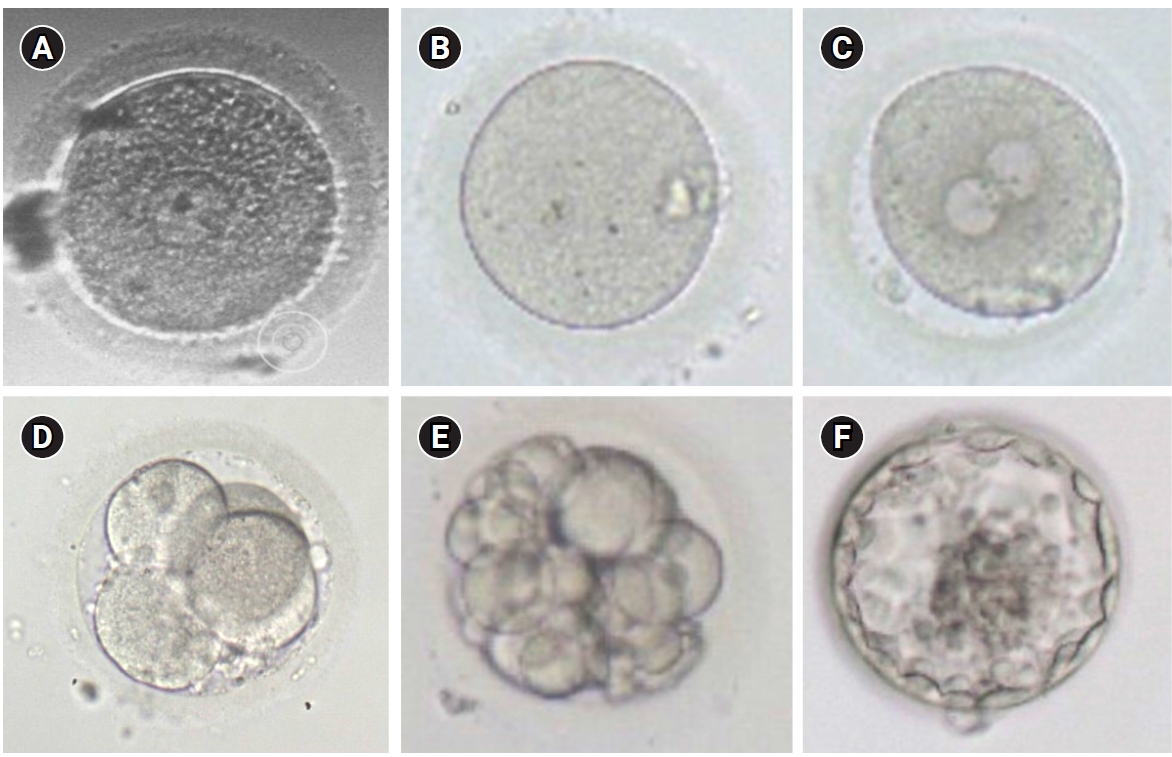
Figure 4 shows the results of the development of human blastocysts from IVM GV oocytes. The present study yielded a total of 13 blastocysts in the experimental group and 4 blastocysts in the control group (Table 5). Furthermore, we observed a statistically significant difference in the number of blastocysts obtained between the experimental and control groups (p<0.05).
Discussion
The complexities around the mechanisms leading to final oocyte maturation remain largely unresolved, which may be why the technique of IVM lacks a precise rationale. The present study was based on the hypothesis that COS can lead to an acceleration of meiotic progression in oocytes, wherein the cytoplasmic maturation often lags behind nuclear maturation [24]. Studies have also shown that the extrusion of the polar body appears much earlier in denuded oocytes; this is deficient in cumulus cells due to the loss of gap junctions, which provide the oocyte with cAMP to maintain dormancy. Therefore, oocytes relieved from the inherent inhibitory effect on meiosis can proceed with nuclear maturation at a faster rate [25]. It has been shown that asynchrony between nuclear and cytoplasmic maturation can cause a negative impact on IVM outcomes. Pre-maturation culture has emerged as a potential method to prolong nuclear maturation and also allows adequate cytoplasmic competence to manifest. Incomplete cytoplasmic maturity can result in multinucleation and aneuploidy, thereby impairing embryo development [8]. However, to date, no consensus exists on the IVM culture system or which, if any, additives are beneficial.
cAMP is the main meiotic inhibitor that keeps oocytes in an arrested state (prophase I) until the hours preceding ovulation. Intra-oocyte cAMP levels are dictated by a balance in the activity of adenylyl cyclase, which assists in the synthesis of cAMP, and PDE, which assists in the degradation of cAMP [26,27]. Furthermore, the cAMP generated in granulosa cells is transported via gap junctions [28,29]. The maintenance of an elevated intra-oocyte cAMP level can keep meiosis in a dormant state. Hence, there is a delay in nuclear maturation, which allows time for cytoplasmic maturation. cAMP levels can be elevated within the oocyte with the help of kinase inhibitors (roscovitine or butyrolactone) [30] or by protein synthesis inhibitors (cycloheximide or puromycin) [31]. As these enzymes can act on both oocytes and granulosa cells, they may impair granulosa cell-oocyte interactions and future embryogenesis. Therefore, in the present study, we used the PDE-3I cilostamide, the action of which is oocyte-specific.
While using cilostamide, the reversibility of meiotic inhibition is an important consideration. A higher concentration of cilostamide can harm the reversibility of meiotic inhibition and spindle structure. Hence, unduly extending the period between transcriptional inactivation and initiation of meiotic maturation may reduce the developmental competence of immature oocytes [20]. Thus, in the present study we used denuded GV oocytes that were exposed to 6 hours of prematuration culture.
As compared to previous studies [32], the present study showed effectiveness in blocking nuclear maturation with 1 μM cilostamide for 6 hours. There was a statistically significant increase in the number of oocytes that remained in the GV stage, as compared to those that progressed towards M1, between the experimental and control groups (92.1% vs. 79.2%, p<0.01). At the end of meiotic arrest, a higher proportion of GV-stage oocytes with the surrounded-nucleolus chromatin configuration [33] indicates a morphological parameter of transcriptional repression. This transformation from a diffused or non-surrounded nucleolus configuration to a compacted or surrounded-nucleolus shape is crucial for successful embryonic development [34]. The results were comparable to those reported in previous studies [20,35]. In our study, we used SAGE medium (Cooper Surgical) to induce oocyte maturation. It has shown proven efficacy for maturing oocytes retrieved from patients with polycystic ovarian syndrome, as well as for the maturation of immature oocytes aspirated prior to treatment of ovarian malignancy [36,37].
The shorter duration of IVM culture is also an independent predictor of better oocyte and embryo quality. The outcome after withdrawal of the inhibitor is still a debatable topic. Son et al. [38] showed that embryos derived from late-maturing oocytes (>48 hours of IVM) were of suboptimal quality in comparison with oocytes maturing between 24 hours and 30 hours of IVM. Therefore, it can be inferred that prolonged blockage of meiotic progression can result in poor-quality embryo outcomes. After the removal of cilostamide blockage, the GV oocytes were cultured in IVM medium and were observed at 26 hours, 36 hours, and 48 hours. A higher number of MII oocytes was observed at 36 hours in the experimental group (25.8%) than in the control group (21.5%). Previous studies have followed several approaches concerning the time of culture for IVM, but there has been no consensus on the optimal maturation time for GV oocytes in IVM cycles in the literature so far [39,40]. We found no significant difference in the fertilization rates among MII oocytes with or without cilostamide exposure (p=0.3). However, the study by Nogueira et al. [32] showed an increased fertilization rate and the study by Jee et al. [20] reported a paradoxical reduction in the fertilization rate.
A previous study by Xie et al. [40] showed that the rates of cleavage and blastocyst development in oocytes maturing at 48 hours after IVM (experimental, 72.2%; control, 19.0%) were significantly lower than in GV oocytes matured 24 hours after IVM (experimental, 91.5%; control, 50.4%). The present study also showed comparable results after 48 hours of IVM culture. From a metabolic perspective, successful IVM of an immature oocyte results from the facilitation of an appropriate environment for the oocyte to attain its developmental competence. COCs require glucose in the medium for metabolism to take place. Cumulus cells possess a substantial capacity for metabolizing glucose, which further produces carboxylic acids and adenosine triphosphate (ATP). The tricarboxylic acid cycle utilizes carboxylic acids within the oocyte. The developmental competence of the oocyte during IVM determines the generation of ATP by the COC. ATP, which is generated by oxidative phosphorylation, also forms the substrate for the production of cAMP. Meiotic inhibition during follicle development is maintained by cAMP. Even in the absence of cumulus cells, immature oocytes can be matured in vitro by the projections of cumulus cells embedded in the zona pellucida. These projections of cumulus cells provide inhibitory and stimulatory signals to the oocytes [41]. Therefore, it is evident that cumulus-free immature human oocytes can mature in IVM medium and also have the potential for early embryonic (cleavage and blastocyst) development.
Furthermore, we observed a statistically insignificant increase in embryo grading in the experimental group, when compared to the control group. However, surprisingly, we found a statistically significant increase in blastocysts in the experimental group (22.8% of MII oocytes injected) as compared to the control group (8.3%). Hence, narrowing the nuclear-cytoplasmic asynchrony with cilostamide plays an instrumental role in obtaining more blastocysts from IVM GV oocytes and could be used as an effective tool in increasing oocyte competence during IVM culture.
In conclusion, the present study provides insights into the IVM culture system for maturing GV oocytes. The developmental competence of oocytes can be enhanced through an improved understanding of the mechanisms that regulate meiotic competence, arrest, and resumption. Further studies are needed to understand the mechanisms through which meiosis is regulated in human oocytes and how oocyte-derived factors influence zygotic genome activation and embryonic developmental competence in human oocytes.