Introduction
Polycystic ovary syndrome (PCOS) is one of the most prevalent gynecological diseases in the world. It has been estimated that 5.6%–6.1% of fertile women around the world are affected by PCOS [1]. Women with PCOS suffer from hyperandrogenism, irregular menstruation, hirsutism, and insulin resistance (IR). IR in these patients is partly due to the mild systemic inflammation that frequently occurs in this syndrome [2]. An increase in proinflammatory cytokines, such as tumor necrosis factor-alpha (TNF-α), can lead to impaired insulin signaling by induction of serine phosphorylation of insulin receptor substrate 1 (IRS1) [3]. One of the main goals of the management of PCOS is reducing IR, which can worsen hyperandrogenism and induce further metabolic abnormalities [4-6].
Several medication strategies have been proposed to reduce IR, for which metformin is one of the most popular recommendations. Metformin enhances glucose homeostasis by inhibiting glucose secretion from hepatocytes and increasing peripheral glucose uptake [7]. Despite the effectiveness of metformin in lowering IR, its side effects such as gastrointestinal distress and interactions with other drugs have raised concerns regarding long-term usage [8]. Some environmental factors (e.g., genetics and nutrition) are also considered as risk factors for PCOS [9-11].
Recent studies have focused on using natural ingredients to reduce the symptoms of PCOS. Active substances, such as isoflavones, which are naturally present in some plants and especially soybean, have been considered as potential alternatives. Genistein, daidzein, and glycitein are the most prominent isoflavones that have been recently studied. Most biological activities of isoflavones are related to their antioxidant and anti-inflammatory properties [12-15]. At the same time, they are known as phytoestrogens, meaning that these compounds have structural similarities with estrogen. Moreover, some beneficial effects may be associated with improvements in insulin sensitivity and amelioration of the lipid profile [16-18].
Studies of animal models have identified favorable results of pure genistein, which could improve the lipid profile and IR [19,20]. Moreover, these changes were accompanied by the promotion of oxidative status and secretion of proinflammatory cytokines [19,21,22]. Romuald et al. [23] found that phytoestrogens supplementation significantly reduced total cholesterol levels and low-density lipoprotein (LDL) in rats. Zhang and Chi [24] investigated the associations of genistein activity with follicle-stimulating hormone receptor and luteinizing hormone receptor expression in rats with PCOS. They found that genistein could improve ovarian function, with increased luteinizing hormone receptor protein expression and decreased follicle-stimulating hormone receptor protein expression.
Genistein supplementation may attenuate the metabolic symptoms of PCOS through its anti-inflammatory and anti-diabetic activities. Therefore, this study aimed to assess the effect of genistein supplementation in comparison to metformin therapy on glycemic indices; the inflammatory, oxidative, and lipid profile; and histopathologic factors in PCOS-induced rats.
Methods
The study was conducted in accordance with the recommendations of the European Council Directive (86/609/EEC) on November 24, 1986, regarding the protection of animals used for experimental purposes (http://data.europa.eu/eli/dir/1986/609/oj). All procedures were approved by the Ethics Committee of Shiraz University of Medical Sciences, Shiraz, Iran (IR.SUMS.REC.1395.S932). Consent for publication was not applicable. Letrozole (Aburaihan Pharmaceutical, Tehran, Iran) was used to induce PCOS (1 mg/kg dissolved in normal saline).
1. Experimental design
In this study, 40 adult (10–12 weeks) female Sprague-Dawley rats were obtained from Shiraz University of Medical Sciences and were housed in a controlled environment (temperature, 22°±2°C; relative humidity, 55%±3%; 12-hour light-dark cycle). All the rats were fed with a normal diet ad libitum. The selected rats had two or three regular estrous cycles during the 2 weeks of vaginal smear observation, and they were in the estrous phase of their reproductive cycle. PCOS was induced using 1 mg/kg of letrozole (Aburaihan Pharmaceutical) dissolved in normal saline. The controls and PCOS-induced rats were evaluated using vaginal smears to determine the irregularity of the estrous cycle and the appearance of persistent vaginal cornification as a symptom of follicular cysts in the ovary.
The rats were initially kept in steel cages for 1 week to acclimate to the new environment. Then, they were randomly divided into four groups (10 rats in each group) as follows: the control group comprised healthy rats, the PCOS group included PCOS-induced rats that received normal saline by gavage, the metformin group comprised PCOS-induced rats treated with 150 mg/kg of metformin (Shafapharma, Tehran, Iran) dissolved in normal saline by gavage [25], and the genistein group included PCOS-induced rats supplemented with 20 mg/kg of genistein (DSM Co., Heerlen, the Netherlands) dissolved in 1% methylcellulose solution. The genistein dosage was determined based on the effective dose of genistein supplementation reported in previous studies [26,27].
2. Study design
This study was carried out for 63 days. The first 21 days were dedicated to induction of PCOS by letrozole and the remaining 42 days to the specific interventions. At the end of the study, the rats were kept fasting overnight and were prepared for sacrifice in the next morning. Ketamine (100 mg/kg) and xylazine (10 mg/kg) were used to anesthetize the rats before cardiocentesis, blood sampling, and ovary sampling. After centrifuging the blood samples at 4,000 rpm for 10 minutes, the serum was separated and stored at –80°C. The ovaries were cleaned of fat, weighed, and fixed in 10% formalin. All rats were weighed on days 1, 21 (after PCOS induction), and 63 (at the end of the study).
3. Assessment of serum biochemical parameters
Fasting blood sugar (FBS), high-density lipoprotein cholesterol, LDL cholesterol, total cholesterol, and triglyceride levels were measured by commercial assay kits (Pars Azmoon, Tehran, Iran) using an Auto-analyzer (BT 1500; Biotecnica, Roma, Italy). In order to estimate superoxide dismutase (SOD) activity and total antioxidant capacity (TAC), enzymatic and colorimetric methods were used based on commercial kit instructions (ZellBio, Lonsee, Germany). In addition, serum malondialdehyde (MDA) was measured using the modified thiobarbituric acid method, as reported by Satoh et al. [28]. Finally, circulating TNF-α levels were determined using the enzyme-linked immunosorbent assay (ELISA) method based on factory instructions of the commercial kit (Diaclone, Besançon, France).
4. Hormonal assay
Regarding to estimate IR, the fasting insulin level was measured using ELISA kits (Mercodia, Sweden). The homeostatic model assessment of insulin resistance (HOMA-IR) was then calculated using the following equation: (fasting insulin[mU/L]×fasting blood glucose [mg/dL])/405.
5. Histopathological examinations
The harvested ovaries were fixed in 10% formalin and firmly embedded in paraffin blocks before being sectioned at 5-µm thickness. Hematoxylin and eosin-stained slides were prepared using the method proposed by Manneras et al. [29]. The slides were then examined for histopathological alterations via a light microscope (Olympus CX31; Olympus, Tokyo, Japan). The histopathological slides were evaluated using different criteria including hyperplasia of theca cells, decreased number of corpora lutea, incomplete luteinization, capsular thickening, and numerous subcapsular follicular cysts, and they were scored as showing no changes, slight changes, or remarkable changes for each criterion.
6. Statistical analysis
The data were reported as mean±standard deviation. The study groups were compared using one-way analysis of variance followed by the Tukey multiple comparison test. All analyses were done using IBM SPSS ver. 19 (IBM Corp., Armonk, NY, USA). The significance level was set at 0.05.
Results
1. Body weight, ovarian weight, and biochemical analysis
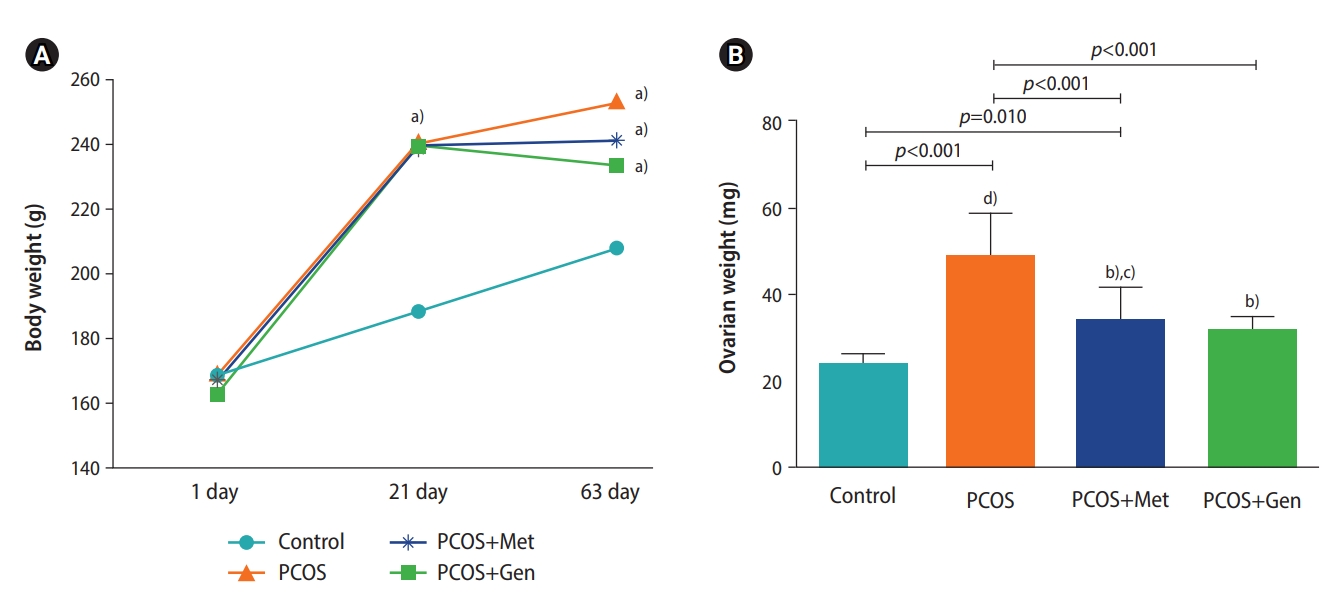
As presented in Figure 1, PCOS induction markedly increased the weight gain in comparison to the healthy group. Accordingly, the weight of the PCOS-induced rats (PCOS, metformin, and genistein groups) was significantly lower than that of the control group on day 21. At the end of the study, the body weight decreased in the metformin and genistein groups in comparison to the PCOS group, but the differences were not statistically significant. Ovarian weights in the PCOS and metformin groups were significantly higher than in the control group. The metformin and genistein groups had significantly lower ovarian weights than the PCOS groups (Figure 1).
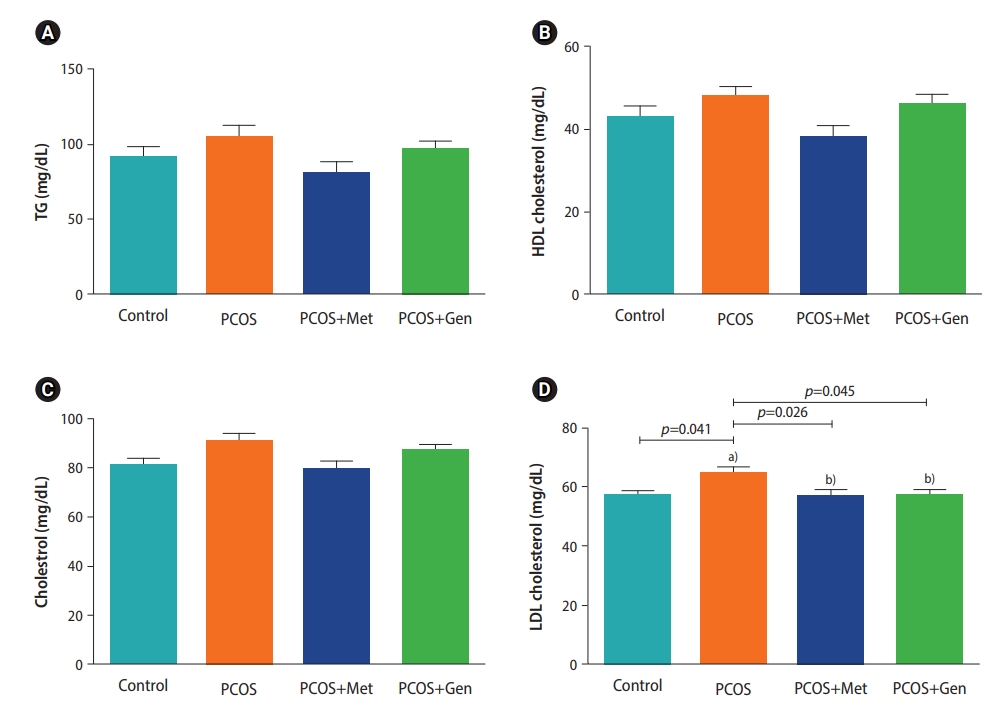
As shown in Table 1, FBS increased significantly in the PCOS group, but not in the rats treated with metformin and genistein (p=0.16). Similarly, fasting serum insulin levels increased only in the PCOS group 43 days after PCOS induction. Thus, PCOS induction was accompanied by an increase in IR in the PCOS group (p=0.012). However, no significant increase was observed in HOMA-IR in the rats receiving metformin and genistein in comparison to the control group (Table 1). The results revealed no significant changes in circulating lipids in the study groups, except for LDL cholesterol, which increased significantly in the PCOS group compared to the control group and significantly decreased in the metformin- and genistein-treated rats compared to the PCOS group (Figure 2).
2. Oxidative and inflammatory indices
The PCOS group had a similar degree of SOD activity to the control group (Figure 3). However, the rats treated with genistein showed a significant increase in SOD activity compared to the healthy rats (p<0.001). Moreover, TAC significantly decreased in the PCOS group compared to the control group, whereas TAC was maintained in both the metformin and genistein groups. In addition, lipid peroxidation as determined by MDA level was significantly elevated in the PCOS group (p=0.01). A similar change was observed in the rats treated with metformin, while no change was observed in the genistein group. Furthermore, PCOS induction did not cause a higher degree of inflammation. Conversely, genistein supplementation reduced the levels of serum proinflammatory cytokines, such as TNF-α (p=0.003), compared to the healthy rats.
3. Histopathological changes
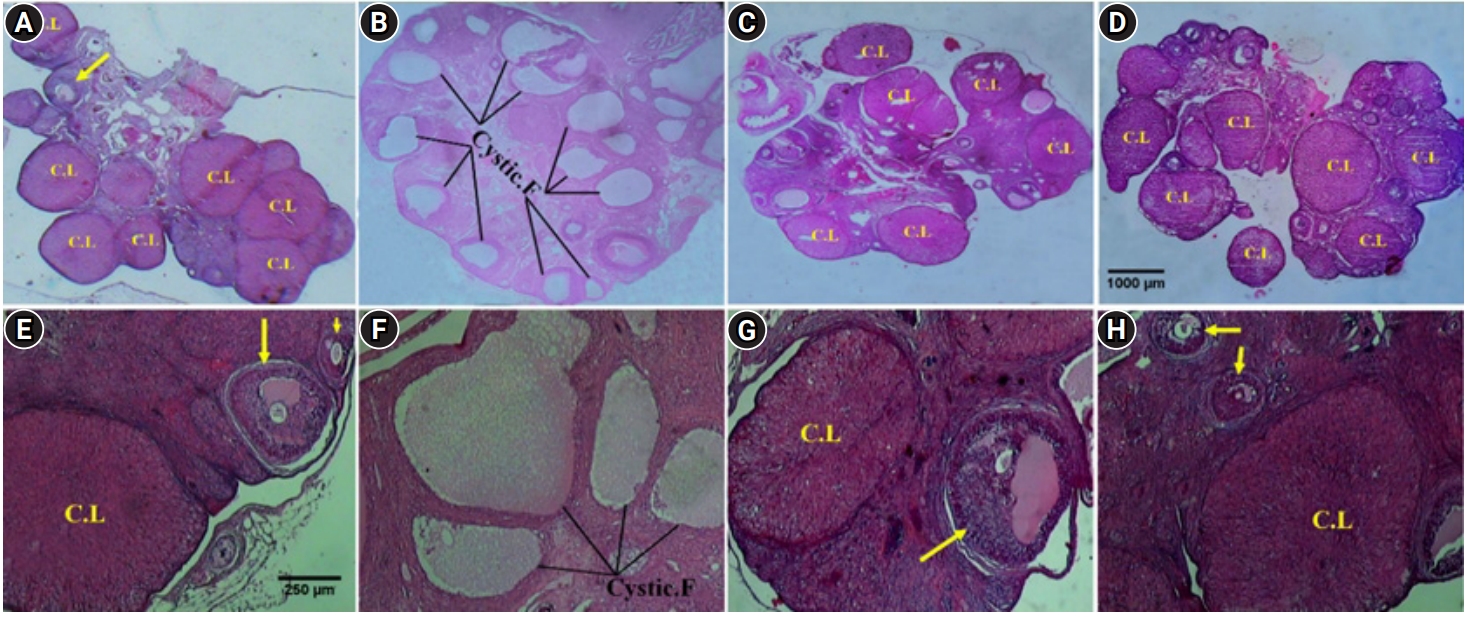
Ovarian sections from the control group showed healthy follicles with oocytes at different developmental stages (Figure 4A and E). Letrozole-treated rats exhibited numerous sub-capsular cysts with very thin or no granulosa layers, and the volume of corpora lutea significantly decreased. In addition, there were few follicles in the ovaries of this group, which were mostly characterized by atresia (Figure 4B and F). Treatment with metformin (Figure 4C and G) and genistein (Figure 4D and H) led to a significant decrease in the number of cysts (p<0.05)and the appearance of healthy follicles and corpora lutea. However, the number of atretic follicles was still significantly higher in these groups than in the control group.
Discussion
The results of the present study identified a greater weight gain in the PCOS-induced groups, which could be attributed to fat accumulation induced by IR, hepatic lipogenesis [30], and the higher level of androgens in PCOS [2]. Treatment with metformin and genistein did not prevent PCOS-induced weight gain, which might be related to the insulin-sensitizing activity of metformin and genistein [31,32].
In the present study, dysregulation of glucose hemostasis and IR in the PCOS group was significantly alleviated after genistein supplementation. It was reported that genistein reduced HOMA-IR mainly by decreasing fasting insulin levels in patients with non-alcoholic fatty liver disease [33]. This effect of genistein has been reported in several diseases [18,33,34]. Genistein may increase glucose uptake by augmentation of shifting glucose transporter type 4 (GLUT4) toward the cell membrane. It has been stated that phosphatidylinositol 3-kinase, protein kinase C, and 5'-adenosine monophosphate-activated protein kinase (AMPK) pathways are involved in the effects of genistein on IR [35].
The histopathological slides in the genistein-supplemented rats showed fewer cysts, and the presence of corpora lutea in the ovaries indicated follicle maturation and ovulation. This aligns with the results of a previous study by Rajan et al. [36], in which treatment of PCOS-induced rats with 100 mg/kg of soy isoflavones caused the development of antral follicles and thickening of granulosa cell layer to the normal state. Interestingly, the glycemic and histopathological changes in the genistein group were fully comparable to those in the metformin-treated group.
Based on oxidative and antioxidant indices, PCOS induction led to higher oxidative status compared to the healthy group. This may itself be a cause of IR induction, as previously reported [37-39]. In addition, folliculogenesis and oocyte maturation were unfavorably affected by oxidative stress [40]. Genistein supplementation in the present study reversed TAC to the normal level and prevented lipid peroxide production by reactive oxygen species, while the metformin group had increased MDA levels. The effect of metformin on increasing TAC may be due to improvements in glucose metabolism, as explained previously. Similarly, genistein can augment antioxidant defenses by upregulating SOD activity. Genistein antioxidant activity mostly relies on two properties; first, the ROS scavenging activity of hydroxyl groups on the 4', 5, and 7 positions [41,42] and second, upregulation of gene expression for antioxidant enzymes (SOD, catalase, and glutathione peroxidase) [43]. The results of the current study confirmed the proposed mechanism, as the genistein group had significantly higher SOD activity than the PCOS and control groups. The current study demonstrated that PCOS induction significantly elevated levels of circulating proinflammatory cytokines, such as TNF-α. This low-grade, but chronic status of inflammation potentially increases the risk of metabolic disorders [44]. It has been reported that increased levels proinflammatory cytokines such as TNF-α in PCOS are independent of body weight. However, the involved signaling pathways are similar, including increased serine phosphorylation, decreased insulin-stimulated IRS1 activation, and downregulation of GLUT4 expression [3,45]. The preservation of normal TNF-α levels in the genistein group may have been due to the anti-inflammatory function of genistein. A possible underlying mechanism could have been the effect of genistein in downregulation of nuclear factor kappa B (NF-κB) expression [46]. NF-κB is a transcription factor involved in the expression of several proinflammatory genes (e.g., TNF-α and interleukin-6) [47]. IR can substantially increase levels of circulating proinflammatory cytokines, which is similar to hyperinsulinemia/hyperandrogenism interaction in PCOS [48].
The present study revealed that no aspects of the lipid profile were affected by letrozole, except for LDL cholesterol levels, which were increased significantly in the PCOS group. Metformin and genistein significantly suppressed this elevation to normal levels. The lipid-lowering action of metformin may be due to activation of hepatocytes via AMPK, which leads to the inhibition of β-Hydroxy β-methylglutaryl-CoA reductase [49]. However, the mechanism of genistein in LDL cholesterol reduction mostly relies on activation of peroxisome proliferator-activated receptor alpha, which subsequently decreases SREBP-2 expression and HMG-CoA reductase activity [50,51].
The promising results in the current study show that genistein may be a possible strategy to confront various complications of PCOS, including IR, dyslipidemia, and oxidative stress. However, further studies are required to investigate the effects of long-term usage of genistein in terms of favorable changes in biological features of PCOS such as hyperandrogenism and to identify its possible side effects. Changes in hormones, especially androgens, should also be examined in future studies.
This study demonstrated that genistein supplementation in rats with letrozole-induced PCOS mitigated IR. Moreover, the histopathological results indicated an increase in luteinization and the development of fewer cysts. These changes were comparable to those observed in the metformin-treated group. Moreover, both genistein and metformin therapy led to lower levels of LDL cholesterol, but only the rats receiving genistein exhibited improvements in antioxidant and anti-inflammatory status. Overall, the results revealed that genistein might be a safe and effective dietary component to ameliorate the complications of PCOS. In addition to acting as a phytoestrogen and regulating metabolic features, genistein exerts antioxidant and anti-inflammatory properties. If these possibilities are confirmed in future studies, genistein may be considered as a supplementary treatment in patients with PCOS.