Introduction
Endometrial fibrosis, the primary pathological feature of intrauterine adhesion, is commonly accompanied by increased deposition and reorganization of extracellular matrix (ECM) proteins, leading to disruptions of endometrial tissue structure, menstrual abnormalities, infertility, and recurrent pregnancy loss [1-5]. Fibrotic progression in the endometrium can be triggered by various factors, including persistent bacterial or viral infections, genetic disorders, or traumatic damage from injury or chemical exposure [6]. Despite the diversity of the causes of endometrial fibrosis, hysteroscopy of fibrotic lesions is only adopted as a current standard surgical resolution for the diagnosis and treatment of this disease [7]. This pathological condition is mostly driven by the interactions of inflammatory cytokines, including interleukin-6, tumor necrosis factor, and transforming growth factor beta (TGF-╬▓), the expression of which is observed in infiltrating inflammatory cells, which tend to migrate towards damaged sites of the endometrium [2,8,9]. However, the specific cellular and molecular mechanisms underlying endometrial fibrosis have yet to be fully elucidated. TGF-╬▓ is known to be a key modulator that facilitates fibrotic progression and the induction of cellular trans-differentiation into myofibroblasts with aberrantly increased ECM deposition [10]. It has been previously reported that TGF-╬▓ is highly expressed in fibrotic tissue in the lung and skin, and conversely, depletion of TGF-╬▓ was found to attenuate experimentally-induced fibrosis progression in the skin, lung, liver and endometrium [11-14]. Binding of TGF-╬▓ to its specific receptor triggers intracellular signal transmission via Smad-dependent and Smad-independent pathways to induce fibrotic progression. The TGF-╬▓ŌĆōinduced expression of alpha-smooth muscle actin (╬▒-SMA) requires Smad3 phosphorylation, which facilitates a remarkable increase in collagen deposition. This process is accompanied by upregulation of the expression of tissue inhibitors of metalloproteinases, which inhibit the activity of ECM-degradative enzymes [15-17]. However, attempts to block the transmission of TGF-╬▓ signaling pathway as candidate anti-fibrotic therapeutic strategies still have limitations due to the characteristics of TGF-╬▓ as a multifunctional physiological mediator. Eupatilin (5,7-dihydroxy-3ŌĆ▓,4ŌĆ▓,6-trimethoxyflavone), which is a major pharmacologically active flavone that has been isolated from Artemisia species, has been reported to suppress TGF-╬▓ receptor signaling by interfering with actin polymerization and acting as a potent inducer of dedifferentiation to reduce fibrotic activity in the liver and lung [18,19]. However, the effects of eupatilin in endometrial fibrosis have not yet been assessed. Since a critical challenge in endometrial fibrotic disease is to understand the progression and prevention of this complex disease entity, it is crucial to identify the cellular and molecular mechanisms underlying endometrial fibrosis. In this study, we present the first report on the impact of eupatilin treatment on TGF-╬▓ŌĆōinduced endometrial fibrosis.
Methods
1. Cell culture and reagents
Ishikawa cells (a well-differentiated human endometrial adenocarcinoma cell line), obtained from American Type Cell Culture (Manassas, VA, USA), were maintained in DulbeccoŌĆÖs Modified Eagle Medium/F12 medium (Gibco, Grand Island, NY, USA) supplemented with 10% fetal bovine serum (Gibco), 100 U/mL penicillin (Gibco), 100 mg/mL streptomycin (Gibco) and 2 mM L-glutamine (Gibco). Recombinant human TGF-╬▓ (cat# L1718; Peprotech, Rocky Hill, NJ, USA) was used at concentrations of 1, 5, or 10 ng/mL. The anti-fibrotic effects of eupatilin (cat# S3846; Selleckchem, Houston, TX, USA) were examined at final concentrations of 25, 50, and 100 ╬╝M. Cells were grown on a Matrigel-coated cover glass (1:8 dilution, growth factor-reduced; Corning, Tewksbury, MA, USA) for further investigation of morphological changes and immunofluorescence (IF) analyses.
2. Immunofluorescence and microscopy
IF staining using an antibody against ╬▒-SMA (1:500; Abcam, Cambridge, UK) was performed as previously described [20].
3. Quantitative real-time reverse-transcription polymerase chain reaction
Total RNA was extracted from Ishikawa cells from three independent experiments using TRIzol reagent (Ambion, Austin, TX, USA). One microgram of RNA was used to synthesize complementary DNA using M-MLV reverse transcriptase (Promega, Madison, WI, USA) and an oligo-dT primer. With one-tenth of the volume of complementary DNA, gene expression was quantitatively analyzed using a reversetranscription polymerase chain reaction (PCR) machine (Bio-Rad, Hercules, CA, USA). Quantitative real-time PCR analysis was performed using FastStart Universal SYBR Green Master Mix (Roche, Basel, Switzerland) on a CFX Connect Real-Time PCR Detection System (Bio-Rad). Amplification consisted of 40 cycles: denaturation at 95┬░C for 10 minutes, annealing at 58┬░CŌĆō60┬░C for 1 minute, and extension at 72┬░C for 1 minute. The primers used in this study are listed in Table 1. The expression levels of genes of interest, which were measured by cycle threshold (Ct) values, were normalized to the Ct values of ╬▓-actin and evaluated as a percentage of the expression level measured in cells of the control group. More specifically, the proportional expression of the gene of interest compared to ╬▓-actin was quantified as 2Ct(gene of interest)ŌĆōCt(╬▓-actin).
4. Statistical analysis
The comparison groups were analyzed using the unpaired Student ttest for parametric distributions. For multiple comparisons, two-way analysis of variance was used, followed by the Sidak multiple comparison test. For all tests, a p-value that was < 0.05 was considered to indicate statistical significance (p< 0.05, p< 0.01, p< 0.001, and p< 0.0001).
Results
1. Eupatilin restores TGF-╬▓ŌĆōinduced fibrotic activity in Ishikawa cells
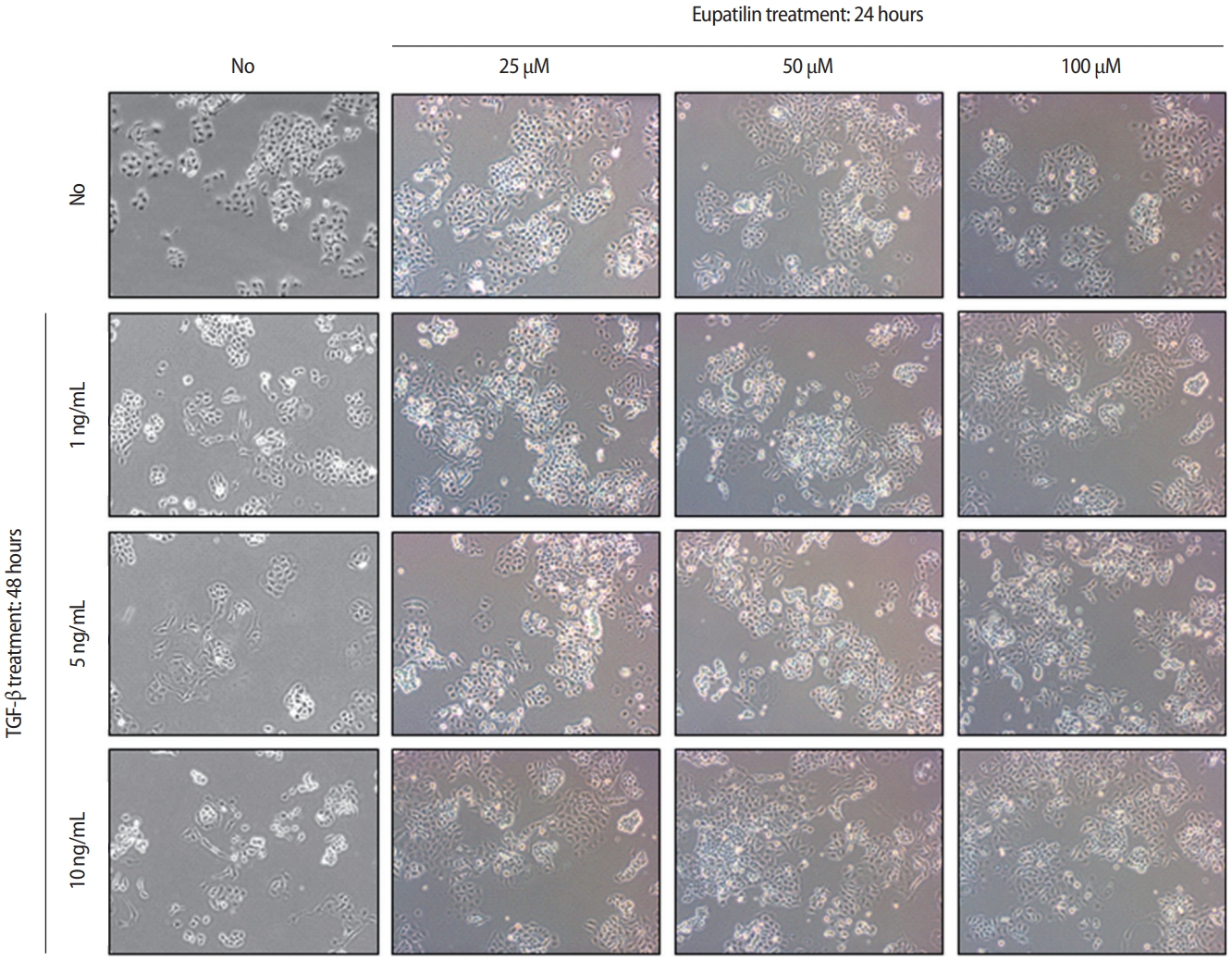
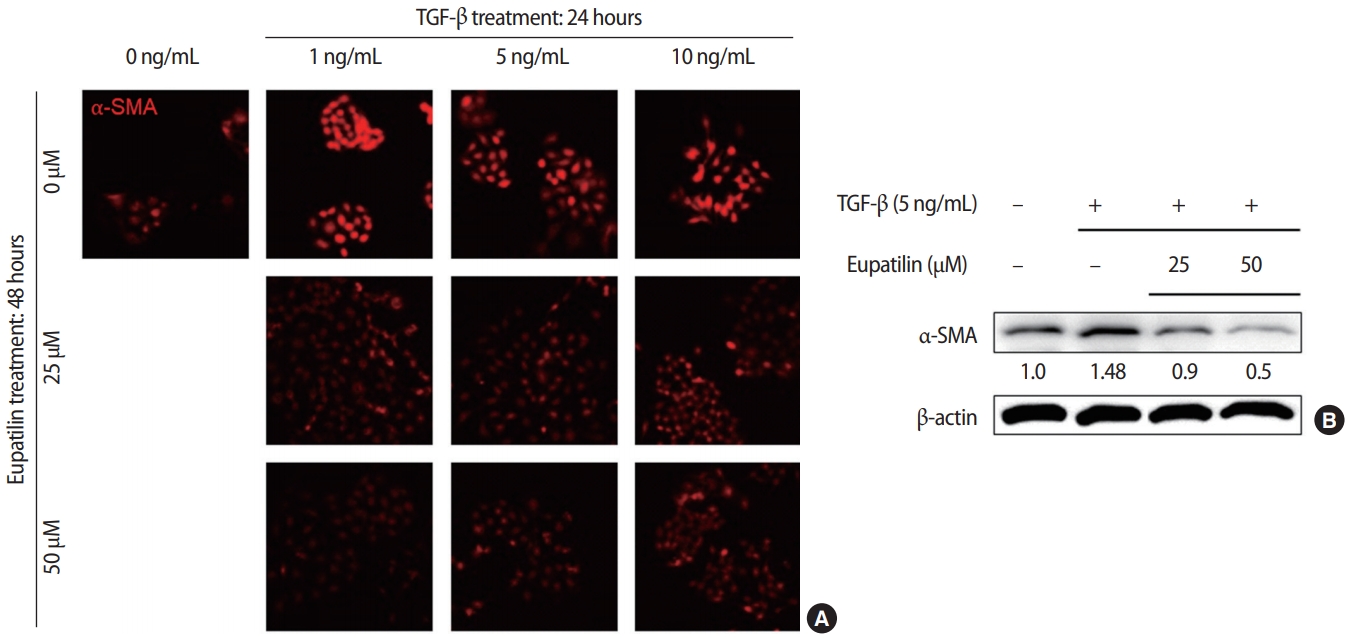
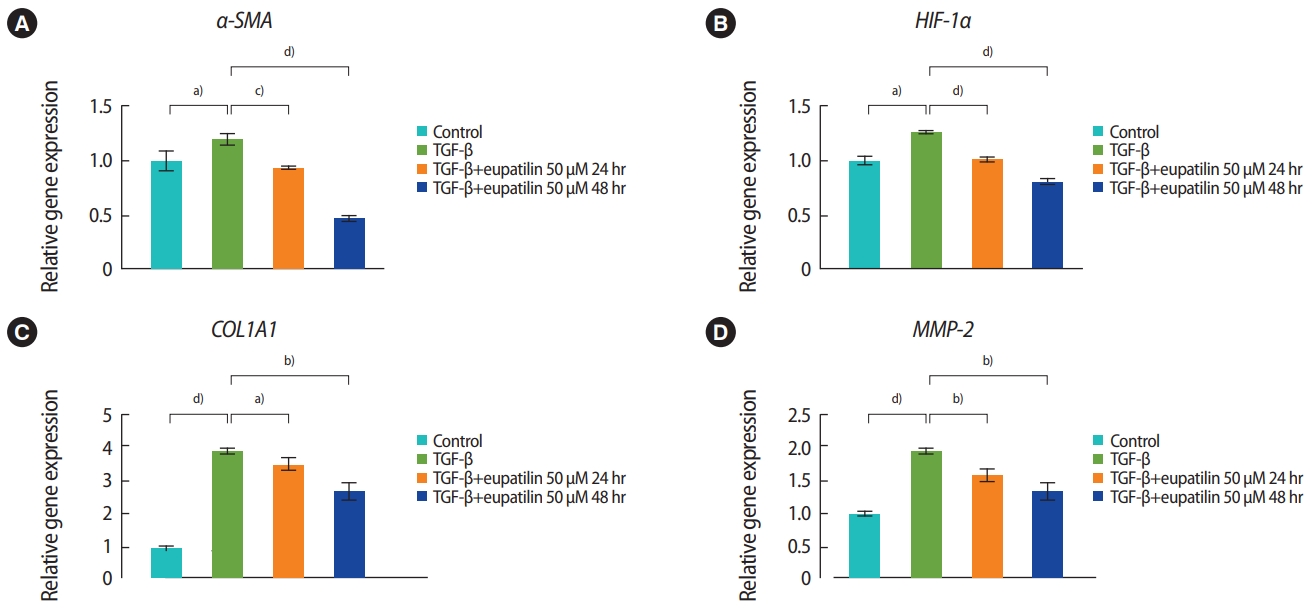
The effects of eupatilin on TGF-╬▓ŌĆōinduced endometrial fibrosis were investigated by assessing morphological alterations in Ishikawa cells. Prior to eupatilin application, the Ishikawa cells were pre-treated with recombinant human TGF-╬▓ for 48 hours to induce fibrosis. As shown in Figure 1, in a dose-dependent manner more fibrotic morphology was observed with higher doses of TGF-╬▓ (far left panel), and additional treatments with eupatilin (25, 50, and 100 ╬╝M) dramatically reduced TGF-╬▓-induced fibrotic phenotypes, as the cells displayed more circular shapes and formed more colonies, which are typical morphological features of Ishikawa cells in culture. IF staining for ╬▒-SMA, a surrogate marker for myofibroblasts, substantiated these findings of morphological alterations. IF and immunoblotting analyses demonstrated that the levels of ╬▒-SMA expression in Ishikawa cells were increased by TGF-╬▓, and subsequent eupatilin treatment drastically inhibited TGF-╬▓ŌĆōinduced ╬▒-SMA to the levels detected in the control group (Figure 2). These data indicate that eupatilin acted directly to attenuate TGF-╬▓ŌĆōinduced fibrotic activity and restored the fibrotic features to the status of normal cells. To determine the efficacy of eupatilin in changing the gene expression of fibrosis markers, we performed quantitative PCR analyses to detect the expression of key genes involved in TGF-╬▓ŌĆōinduced endometrial fibrosis in Ishikawa cells and then measured the changes in their expression levels in response to eupatilin treatment. ╬▒-SMA, hypoxia-inducible factor 1 alpha (HIF-1╬▒), collagen type I alpha 1 chain (COL1A1), and matrix metalloproteinase-2 (MMP-2) were highly upregulated by 48 hours of treatment with TGF-╬▓, and their expression was significantly reduced by treatment with 50 ╬╝M eupatilin (Figure 3). Longer exposure to eupatilin resulted in a greater reduction of the expression of all four of those genes. Of note, both COL1A1 and MMP-2 failed to recover to the levels of the control cells with 25 ╬╝M eupatilin treatment (data not shown), implying that certain levels of eupatilin might be required to overcome the fibrotic effects induced by TGF-╬▓. Taken together, these data reveal that TGF-╬▓ŌĆōinduced phenotypic and molecular alterations in endometrial fibrosis were both restored by eupatilin treatment, suggesting this drug as a potent therapeutic intervention for patients with fibrosis in the endometrium.
Discussion
In recent years, autologous transplantation of bone marrow-derived stem cells [21] and menstrual blood-derived stromal cells [22] has been performed in patients with endometrial fibrosis or intrauterine adhesion. Cell therapies targeting endometrial fibrosis might be an option for stimulating the growth of the endometrium and recovering reproductive function [21,22]. However, the molecularbased mechanisms underlying anti-fibrotic activity have not yet been fully elucidated. Previous studies have suggested that microRNAs (miRNAs) might serve as potential therapeutic targets for endometrial fibrosis. Xiao et al. [23] demonstrated that miRNA-340 induced by bone marrow mesenchymal stem cell treatment stimulated the regeneration of rat endometrial tissue subjected to mechanical damage. Furthermore, Ning et al. [2] reported that down-regulation of miRNA-326 was strongly associated with intrauterine adhesion, characterized by endometrial fibrosis and high levels of TGF-╬▓1, ╬▒-SMA, COL1A1, and fibronectin, as we observed in the present study. Recently, two small molecules (pirfenidone and nintedanib) were approved for idiopathic pulmonary disease, as they showed combinatorial anti-inflammatory, antioxidant, and antifibrotic effects [24,25]. Moreover, Kim et al. [18] reported that eupatilin treatment attenuated the progression of bleomycin-induced lung fibrosis and TGF-╬▓ŌĆōinduced fibrotic activity in hepatic stellate cells by disturbing Smad3 phosphorylation, which subsequently led to myofibroblast dedifferentiation and reversal of fibrotic progression. They demonstrated the effects of eupatilin on actin depolymerization and suppression of induction of the epithelial-mesenchymal transition [18]. However, no report has yet provided evidence for the impact of eupatilin in endometrial fibrosis.
The present study demonstrates that eupatilin treatment restored TGF-╬▓ŌĆōinduced fibrotic activity in endometrial epithelial cells, as shown by a reduction in the degree of fibrotic phenotypes and a more circular shape of individual cells. This conclusion is also supported by the significant observed reduction in the expression of ╬▒-SMA, COL1A1, and MMP-2, which were strongly induced by TGF-╬▓, by eupatilin treatment. Due to technical obstacles and limitations, we evaluated the effects of eupatilin treatment on fibrosis that was artificially induced by TGF-╬▓ in Ishikawa cells in vitro instead of using primary endometrial epithelial cells. Additionally, Ishikawa cells only represent the epithelial compartment of the endometrium. Since fibrotic activity and its progression result from communication between the endometrial epithelium and stroma, our future research will address the relevance of eupatilin treatment for fibrotic activity in endometrial stromal cells in association with the endometrial epithelial layer. In conclusion, the findings of the present study suggest that suppression of TGF-╬▓ŌĆōinduced signaling by eupatilin might be an effective therapeutic strategy for the treatment of endometrial fibrosis. Preventing endometrial fibrosis by understanding its fundamental molecular mechanisms may potentially provide therapeutic strategies for converting fibrotic endometrial tissue into functionally active endometrium.