Introduction
Various techniques have been introduced to satisfy the demand for suitable mammalian oocyte and embryo culture systems. For example, a co-culture system with somatic cells such as cumulus cells could increase the fertilization and blastocyst formation rate [1,2,3]. The addition of growth factors or cytokines that are secreted by reproductive organs to the culture medium also could increase embryonic development and implantation rate [4,5]. In fact, insulin-like growth factor-I (IGF-I) and IGF-II increase formation of blastocysts and their cell number, especially increasing the cell number of inner cell mass (ICM) [6,7], while leukemia inhibitory factor (LIF) increases blastocyst formation and trophoblast outgrowth [8]. However, even the usefulness of upgrading culture systems and the precise mechanisms of those materials in oocytes or embryos remain unclear. Furthermore, their relatively low stability should be overcome for general use in long-term culture systems.
Estrogen-related receptor β (Esrrb) is a member of the orphan nuclear receptors. It is known to be a regulator of the expression of pluripotency-related genes such as Oct4 and Nanog, and maintains the undifferentiation and pluripotency of stem cells [9]. Oct4 also maintains the formation of the ICM, and it regulates blastocyst differentiation and embryonic development during peri-implantation [10]. Additionally, Esrrb plays an essential role in adequate placental development. In fact, knockdown of Esrrb induces differentiation of embryonic stem (ES) cells [11] and Esrrb-null mutant mice die at 10.5 days post conception due to placenta abnormality [12].
In a previous study, recombinant protein conjugated with cell penetrating peptide (CPP) using 7X-arginine (R7) was translocated into human mesenchymal stem cells or ES cells efficiently with biological activity [13,14]. In the present study, we have analyzed the in vitro development of mouse embryos by delivery of CPP-ESRRB recombinant protein into a culture system and developed a novel embryo culture system using exogenous protein delivery.
Methods
1. Animals and ovarian hyperstimulation
Four- to 5-week-old female and 8- to 10-week-old male B6D2F1 mice (Samtako, Seoul, Korea) were kept under controlled light and temperature conditions with free access to water and food under 12 hours light and 12 hours dark. All animal experiments were approved by the Animal Care and Use Committee of CHA University. The female mice were superovulated via intraperitoneal injection of 5 IU pregnant mare serum gonadotropin (PMSG, Sigma-Aldrich, St. Louis, MO, USA), followed by injection with 5 IU hCG after 48 hours (Sigma-Aldrich). The female mice were then mated with the male mice and their vaginal plugs were checked at 12 hours after mating.
2. Collection and culture of mouse embryos or oocytes
Two-cell embryos were collected from the oviduct at 46 hours after hCG injection and cultured in potassium simple optimization medium amino acid (KSOM, Millipore, Billerica, MA, USA) under mineral oil (Sage, Trumbull, CT, USA) in an incubator at 37℃ and 5% CO2 in air. Then, 8-cell embryos derived in vitro were cultured in KSOM with or without 2 µg/mL of CPP-conjugated ESRRB protein for 48 hours. At 120 hours after hCG injection, in vivo-derived blastocysts were recovered from the uterus and used as a control group.
Germinal vesicle-stage (GV) oocytes were collected from the ovaries at 48 hours after PMSG injection. Metaphase II (MII)-stage oocytes were collected after in vitro maturation of GV oocytes in maturation medium (KSOM containing 0.4% bovine serum albumin (BSA, Sigma-Aldrich) for 12 hours.
3. Construction of a CPP-ESRRB expression vector and purification of recombinant CPP-ESRRB protein
In order to link Esrrb to 7R (a CPP domain) and a 6 histidine (His)-tag, mEsrrb cDNA was inserted into a pET20b(+) vector to construct an expression plasmid according to the protocol of Jo et al. [15]. The CPP-ESRRB protein expression vector was transformed into BL21 (DE3) pLysS-competent cells (Stratagene Inc., La Jolla, CA, USA). Transformed cells were cultured in lysogeny broth (LB) medium with ampicillin and grown at 37℃ on a shaking plate. The protein synthesis was induced with 1 mM isopropyl-1-thio-β-D-galactopyranoside (IPTG, Sigma-Aldrich) and cell cultures were further grown at 20℃ for 8 hours. The cells were lysed for 30 minutes at room temperature in a lysis buffer with NP-10 (50 mM NaH2PO4, Sigma-Aldrich); 300 mM NaCl (Sigma-Aldrich); and 10 mM imidazole (Sigma-Aldrich), pH 8.0; 1 mL of lysozyme (Sigma-Aldrich) solution; and benzonase (3 units/mL; Qiagen Inc., Basel, Switzerland). The lysate was then separated into supernatant and lysate pellets were obtained by centrifugation at 15,000 rpm. The soluble supernatant was purified using Ni-NTA Superflow Columns (Qiagen Inc.). The purified CPP-ESRRB protein was mixed with 2× Laemmli sample buffer and boiled for 5 minutes for 15% SDS polyacrylamide gel electrophoresis (SDS-PAGE). Gels were stained with Coomassie Brilliant Blue R250 for visualization of the protein. The purified CPP-ESRRB protein was also confirmed by western blotting using ESRRB antibody.
4. Counting the cell numbers of the blastocysts
The number of ICM and trophectoderm (TE) cells in the blastocysts derived from embryo experiments were counted by differential staining using two chromatin-specific fluorochromes with different fluorescent spectra: propidium iodide (Sigma-Aldrich), which enters only cells with damaged membranes, and bisbenzimide (Hoechst 33342, Sigma-Aldrich), which passes through both damaged and intact membranes. At day 5, the zona pellucida (ZP) of the collected blastocysts was removed by brief exposure to acid Tyrode's solution (Sigma-Aldrich). The ZP-free embryos were exposed to a 1:5 dilution of whole rabbit anti-mouse serum (Sigma-Aldrich) for 1 hour and washed three times with DPBS (Hyclone) containing 0.1% BSA for 5 minutes each. Then, the embryos were placed into a 1:10 dilution of guinea pig complement (Sigma-Aldrich) for 1 hour. Propidium iodide and bisbenzimide were added to the complement solution to a final concentration of 10 µg/mL and 10 µg/mL, respectively. Then, they were briefly washed in DPBS (Hyclone) containing 0.1% BSA (Sigma-Aldrich) and were mounted between the slide and coverslip and examined under ultraviolet light using an Axio Imager A2 microscope (Carl Zeiss, Jena, Germany) fitted for epifluorescence. The nuclei of the ICM were labeled with bisbenzimide and appeared blue, while the nuclei of the TE cells appeared pink.
5. RNA extraction, reverse transcription- and quantitative real-time PCR
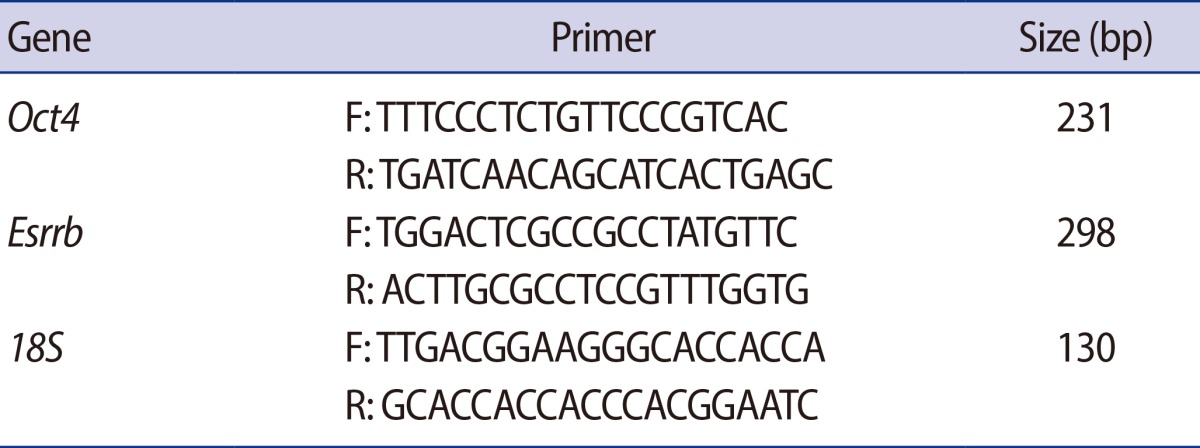
Total cellular RNA was extracted from 15 GV oocytes and embryos using the TRIzol reagent (Invitrogen, Carlsbad, CA, USA). The first strand of cDNA was synthesized using a PrimeScript 1st strand cDNA Synthesis kit (Takara Bio, Shiga, Japan) according to the manufacturer's instructions. Reverse transcription polymerase chain reaction (PCR) was conducted using G-Tag (Cosmogene Tech, Seoul, Korea) and specific primer sets (Table 1). The cycle conditions were as follows: 94℃ for 5 minutes, followed by 35 cycles of a 94℃ denaturation period for 30 seconds, a 60℃ annealing period for 30 seconds, and a 72℃ elongation period for 40 seconds, with a final elongation period at 72℃ for 10 minutes. All PCR products were separated by 2% agarose gel electrophoresis. For quantification of the expression level, real-time PCR was performed by using iQTm SYBR Green super mix (Bio-Rad Laboratories, Hercules, CA, USA) on a Bio-Rad iQ5 real-time PCR machine. Each gene was normalized with 18S as housekeeping control. The results were analyzed by using the delta-delta Ct method with use of the 18S normalization control.
6. Western blot analysis
Western blot analysis was performed to characterize the CPP-ESRRB and confirm delivery of CPP-ESRRB protein into the embryos. Each sample containing 60 embryos, which were separated by 12% SDS-PAGE; after that, they were transferred onto polyvinylidene difluoride membranes (Bio-Rad Laboratories) stored for 90 minutes. Blocking was performed in DPBS containing 3% BSA and 0.1% Tween-20 (Sigma-Aldrich) for 2 hours at room temperature. The membranes were incubated in blocking solution with anti-ESRRB (Santa Cruz Biotech-nology, Santa Cruz, CA, USA, 1:200) or anti-His-tag (Sigma-Aldrich, 1:700) overnight, and then each sample was evaluated with a horseradish peroxidase-conjugated secondary antibody (Santa Cruz Biotechnology) diluted to 1:2,000 with DPBS containing 0.1% Tween-20 (Sigma-Aldrich) for 1 hour. Immunoreactivity was detected using Western Blotting Luminol Reagent (Santa Cruz Biotechnology) and developed on Hyperfilm ECL X-ray film (Amersham Pharmacia Biotech, Buckinghamshire, UK).
7. Immunocytochemistry
Embryos cultured with or without CPP-ESRRB for 24 hours and 48 hours were fixed in 4% paraformaldehyde with 0.1% BSA (Sigma-Aldrich) at 4℃ for 30 minutes, and washed twice with DPBS (Hyclone) containing 0.1% BSA. After that, the embryos were permeabilized with 0.1% Triton X-100 (Sigma Aldrich) for 20 minutes and blocked in DPBS (Hyclone) with 3% BSA and 0.05% Tween-20 (Sigma-Aldrich) for 2 hours at 4℃. They were then stained overnight at 4℃ with primary antibodies anti-His-taq (1:100, mouse polyclonal, Sigma-Aldrich) or anti-ESRRB (1:100, rabbit polyclonal, H70, Santa Cruz), followed by 2 hours incubation at 4℃ with Alexa Fluor 555-labeled goat anti-rabbit, 488-labeled goat anti-mouse secondary antibodies (Molecular Probes, Eugene, OR, USA) diluted to 1:200 with 3% BSA in DPBS (Hyclone). All of the samples were counterstained with 1 µg/mL 4, 6-diamidino 2-phenyindiol (DAPI, Sigma Aldrich) diluted to 1:1000 with DPBS for 15 minutes at room temperature. All sample images were captured with an LSM 510 META confocal laser-scanning microscope (Carl Zeiss). The stained cell images were reconstructed using LSM 510 META software.
Results
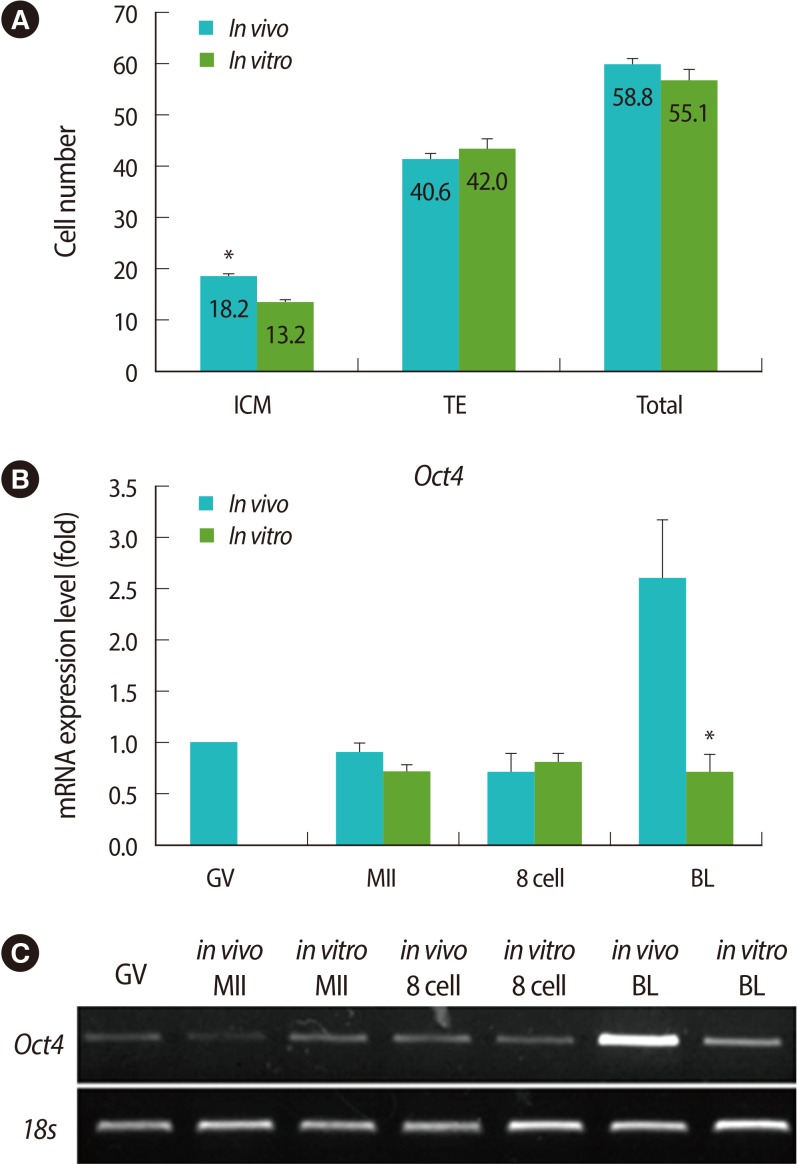
1. Comparison of cell numbers and Oct4 mRNA expression in blastocysts derived in vivo and in vitro
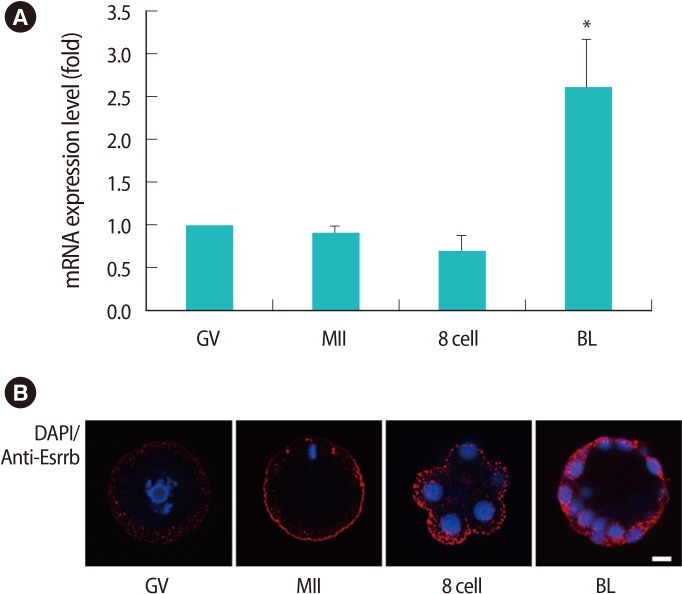
To verify the cell numbers of blastocysts derived from the groups cultured in vitro and in vivo, their cell numbers fo TE and ICM were counted using differential staining. The average cell number of ICM was lower in in vitro-derived blastocysts than in vivo-derived ones (Figure 1A). In addition, endogenous expression levels of Oct4, which regulates ICM formation, was also analyzed in the oocytes and embryos of each stage by real-time PCR. Like the cell number of ICM, the mRNA expression of Oct4 was lower in the in vitro-derived blastocysts than the in vivo-derived ones (Figure 1B, C).
3. Characterization of CPP-ESRRB recombinant protein
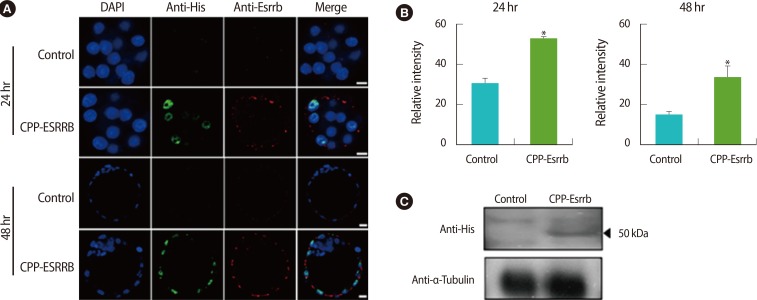
The expression of CPP-ESRRB protein was induced by IPTG and was purified using His-tag at the C-terminal. The purified CPP-ESRRB protein was characterized through electrophoresis using SDS-PAGE gel and staining with Coomassie Brilliant blue R250 (Figure 3A). This result was reconfirmed by Western blot analysis using ESRRB-specific antibody (Figure 3B). A major single band was displayed, and that was CPP-ESRRB in 50 kDa (Figure 3B Lanes 2-4). A lower band was assumed to represent an additional protein, non-purified E. coli protein, or truncated CPP-ESRRB. In contrast, E. coli cell extracts containing the pET-empty construct did not display the major single band representing CPP-ESRRB-His (Figure 3A Lane 1, Figure 3B Lane 1). Figure 3 shows the high purity of the ESRRB isolation identified in this experiment.
4. Delivery of CPP-ESRRB into the embryo
To observe the delivery of CPP-ESRRB recombinant protein into embryos during in vitro culture, we performed immunocytochemistry and western blot analysis. In immunocytochemical analysis using anti-His-tag and anti-ESRRB antibodies, the CPP-ESRRB delivered into the embryos was detected in their cytoplasm and nucleus at 24 hours and 48 hours after treatment, respectively (Figure 4A). However, their intensity was high at 24 hours after treatment and then slightly decreased at 48 hours (Figure 4B). Western blot analysis using anti-His-tag antibody also clearly showed that CPP-ESRRB recombinant protein was delivered in embryos of the treated group (Figure 4C).
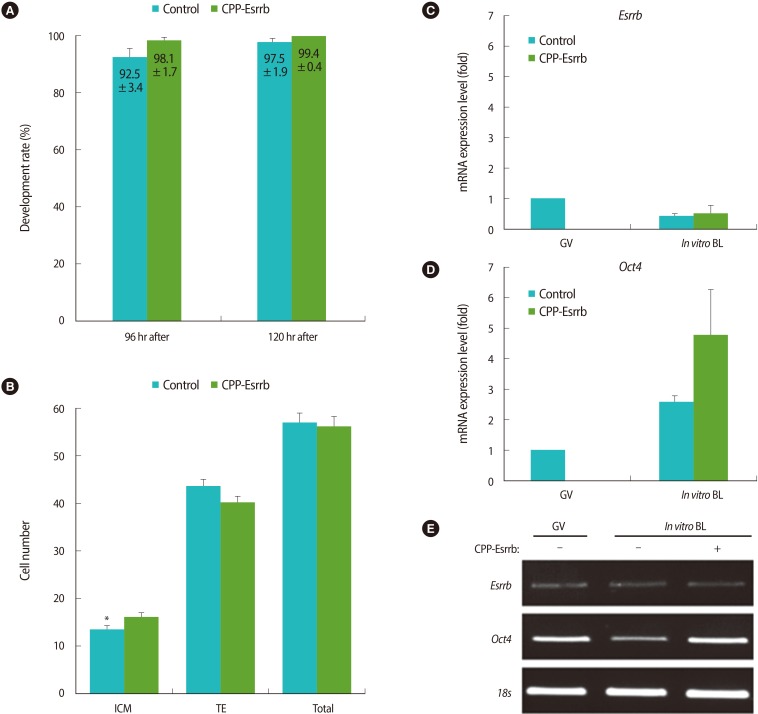
5. Effects on further development of embryos treated with CPP-ESRRB during in vitro cultivation
As shown in the Figure 5A, treatment of CPP-ESRRB during in vitro culture was either performed or not; there were no differences in the rates of blastocyst formation between the two groups (98.1±1.7% vs. 92.5±3.4% at 96 hours after hCG injection and 99.4±0.4% vs. 97.5±1.9% at 120 hours after hCG injection, respectively; p>0.05). However, the cell number of ICM in the CPP-ESRRB-treated embryos was greater than in the non-treated embryos (16.0±1.1% vs. 13.4±0.9%; p<0.05) (Figure 5B). Interestingly, there were no significant differences in the cell number of total and TE in blastocysts obtained from both groups.
6. Effects on the Oct4 expression in embryos treated with CPP-ESRRB during in vitro cultivation
After the embryos were cultured in KSOM with or without 2 µg/mL of CPP-ESRRB for 48 hours, in vitro grown blastocysts were collected from both groups and the expression of Esrrb and Oct4 mRNAs were analyzed by real-time PCR. As shown in Figure 5C, the expression level of endogenous Esrrb did not differ between the treated group and non-treated groups. However, blastocysts in the CPP-ESRRB-treated group have shown a higher expression of Oct4 mRNA compared to those of the non-treated group (Figure 5D, E).
Discussion
In vitro culture systems for embryonic development are indispensable for performing assisted reproductive technology (ART) such as IVF, ICSI, and cryopreservation. However, the manipulation of embryos or oocytes under suboptimal conditions could cause abnormal development of the fetus and placenta [16]. Also, in fact, the cell number of blastocysts derived in vitro was lower than from those in vivo [17]. In the present study, we have found that the cell number of ICM and Oct4 mRNA expression of CPP-ESRRB-treated embryos was greater than those of non-treated embryos. Although we cannot completely exclude the possibility that other factors in the culture medium have affected embryonic development, we have shown that the addition of CPP-ESRRB to culture medium improved the quality of embryos in vitro.
According to recent reports, there are three mRNA alternative splicing isoforms of human Esrrb. The short form is located in the nucleus and activates transcription. The short form of human ESRRB is the actual human ortholog of mouse Esrrb [18]. ESRRB protein also has cysteine-rich domains that can combine with DNA for the regulation of transcription [19], and Esrrb was expressed within the nuclei of epithelial, stromal, immune, and endothelial cells [20]. Moreover, Esrrb regulates the expression of Oct4, and Oct4 was known to regulate Nanog as well, so these genes were regulated by each other [21]. In fact, Oct4-deficient embryos can develop to the blastocyst stage, but their ICM are not pluripotent [22]. Instead, they are restricted to differentiation along the extra embryonic trophoblast lineage. Therefore, Oct4 and Nanog are both required in vivo and in vitro for establishment and maintenance of pluripotent ICM in mouse blastocysts [23].
In the present study, we used a CPP protein delivery system known as protein transduction domains or membrane transduction peptides. They have been thought to be a useful tool due to their ability to translocate across cellular membranes [24]. They consist of short sequences of amino acids, such as arginine or lysine residues, which confer a positive charge to the CPPs [25]. We have chosen oligoarginine (R7), which does not enter the cell by endocytosis. Also, it can not only pass through the cell membrane easily, but also move throughout the nucleus [26]. As shown in Figure 4, we have found that CPP-conjugated 50 kDa ESRRB can successfully penetrate the ZP and cytoplasm of the embryo and move into the nucleus. In addition, treatment with CPP-ESRRB has not shown a negative effect on embryonic development, but can increase ICM formation during mouse embryogenesis (Figure 5). This suggests that CPP-ESRRB could manipulate some embryonic processes in other mammalian species that show poor ICM development under suboptimal culture conditions or after harsh ART procedures.
In conclusion, recombinant CPP-ESRRB could be easily delivered into embryos by CPP and may increase the level of Oct4 mRNA and the rate of ICM formation. This technology could also be a useful tool for improving culture systems in vitro. In further research, we should observe the implantation rate, full-term development, and second generation to confirm the stability of the culture system using CPP-ESRRB.