Introduction
Self-renewal activity and the ability to differentiate into various functional cells are two major characteristics of stem cells [1]. Stem cells have thus been considered among the therapeutic biomaterials of use in regenerative medicine, which can restore the function of damaged cells, tissues, or organs. Before clinical application, however, a number of difficulties in manipulating stem cells should be overcome, and a variety of alternative technologies have been suggested to control stem cell fate and function.
Recent studies have demonstrated that conventional stem cell technologies do not meet the minimal requirements to precisely regulate stem cell function [2]. To overcome this limitation, several scientists have taken an interest in regulating stem cells under a specific microenvironment, which uses artificial extracellular matrices (ECMs) to control the stem cell self-renewal and differentiation activity [3]. The use of artificial ECMs for creating three-dimensional (3D) microenvironments would yield several benefits, mimicking the in vivo microenvironment of the undifferentiated or differentiated period at both the cell and tissue levels [4]. Biophysical microenvironments, including the stiffness of the substrate, nanotopography of the adhesion surface, microgeometric forces, and extracellular forces, must be constructed prior to the use of this 3D system, and the influence of stem cell fate in vitro through this artificial microenvironment can efficiently prevent the direct genetic manipulation of stem cells, which would otherwise limit the feasibility of clinical application [5-8]. Having access to artificial ECMs thus accelerates niche-related studies on manipulating stem cell self-renewal and differentiation; the data obtained from 3D cultures can be comparable to those obtained from conventional two-dimensional (2D) cultures.
Application of 2D niche systems to stem cell self-renewal maintenance
As an initial step, most studies have employed embryonic stem cells (ESCs) in both experimental animals and humans. However, data from the culture of stem cells under conventional 2D systems provide a wealth of information that should assist in the development of innovative 3D culture systems. Therefore, careful consideration of stem cell culture in 2D systems is a prerequisite for developing 3D culture systems.
1. Cellular niche systems
ESCs have generally been co-cultured with feeder cells such as xenogenic embryonic fibroblasts to maintain self-renewal activity. The feeder cells used for stem cell culture are able to supply growth factors, cytokines, and other extracellular matrix components such as leukemia inhibitory factor (LIF), activin, Wnt, bone morphogenetic proteins (BMPs), insulin-like growth factor (IGF), laminin, and vitronectin for maintaining the undifferentiated state of ESCs [9,10], which can create a suitable 2D environment. When applying these 2D systems to human ESCs, animal-derived feeder cells can cause unexpected disadvantages such as uncertain data outcomes and xenotransmission of unknown pathogens [11]. To avoid these handicaps, studies on the development of feeder-free cultures and defined culture systems have been strongly encouraged [12].
Since the initial studies of Richards et al. [13], human feeder cells such as neonatal foreskin fibroblasts [14], fetal muscle, fetal skin, adult fallopian tube epithelial cells [13], adult muscle, adult skin [15], marrow-derived stromal cells [16], amniotic fluid fibroblasts [17], placenta-derived fibroblasts [18], and human ESC (hESC)-derived fibroblasts [19] have been employed to provide a suitable cellular niche for hESCs without the use of xenogenic cells. A positive outcome of the replacement was reported [15], although batch differences in feeder cell-based culture methods is considered another uncertainty that impedes the establishment of stable culture conditions. To further develop defined stem cell culture systems, the use of non-cellular niches can subsequently be considered.
2. Non-cellular niche systems
An initial attempt has been made to employ non-cellular niches for the development of defined ESC culture systems, and artificial ECMs were thus employed. Instead of feeder cells, Xu et al. [12] used Matrigel-coated dishes. ESCs were successfully cultured with the use of fibroblast-conditioned medium, to which co-culture technology was applied for the culture of other stem cell lines [20]. Each medium conditioned with different cells had a different capacity to maintain ESC self-renewal [21]. However, some showed negative results in attempts to support the long-term culture of ESCs, which demonstrates their unsuitability as a standardized culture regime [22]. Such a limitation directly encourages the development of a defined culture medium and the refinement of ESCs for suitable ESC culture methods.
Amit et al. [23] first reported the successful use of knockout serum for ESC culture as a replacement for bovine serum, although this semi-defined serum replacement contains xenogenic or undefined substances such as bovine serum albumin [24]. Subsequently, a culture medium containing human-originated recombinant proteins was developed (X-VIVO 10 medium). As suitable supplements for hESC culture, basic fibroblast growth factor (b-FGF), stem cell factor (SCF), recombinant human FMS-like tyrosine kinase 3 ligand (rhFlt3L), and LIF [25] were recommended. TeSR1 using recombinant proteins and purified material from humans was also suggested as a culture supplement for hESC culture [26]. Discovery of the alternative material Matrigel was developed and the single use of laminin, one of the Matrigel components, was successful for the culture of undifferentiated hESCs [16]. It has also been reported that laminin isoforms 111, 332, and 511, vitronectin, and E-cadherin support the adhesion and proliferation of hESCs [27-29].
Application of 3D non-cellular niche systems for stem cell self-renewal
1. Biomaterial-based 3D scaffold system
Among 3D scaffolds being constructed with various biocompatible biomaterials, hydrogel-based ECMs have been employed for the culture of stem cells. The hydrogel is a cell-friendly, 3D macromolecule platform formed by the crosslinking of hydrophilic polymers, which can absorb H2O molecules to over five hundred times its own weight. In response to the salt concentration, pH, temperature, and crosslinking method, the chemical structure of the hydrogel can change in unlimited ways, so many biomedical [30] and pharmaceutical [31] applications may be possible, such as drug delivery [30], artificial tissue [32], membrane fabrication [33], and matrix construction [34].
To date, natural material-based hydrogel systems have been developed. Hydrogel-based, protein-like materials such as Matrigel [35,36], collagen [37], fibrin [38], and silk [39,40], polysaccharide-like materials such as hyaluronic acid [41,42], alginate [43,44], dextran [30], chitosan [45,46], and agarose [47], and DNA [48] have been used. In most of these cases, however, difficulties with manipulating their mechanical properties, the degradation rate, and the reproducibility occurred due to their undefined protein composition, which limited the modification of binding domains or sites in cellular activity and immune responses. To overcome this limitation, a hydrogel-based, synthetic (chemically defined) material has subsequently been suggested, which has a number of advantages including easy manipulation of mechanical properties, degradation, shape, and reproducibility. Many biological processes can be regulated by the use of synthetic hydrogel components, which further makes it possible to develop transplantable biomaterials. Hydrogels polymerized with peptide [49] or poly (lactic-co-glycolic acid) (PLGA) [50] have subsequently been developed, and recently, the use of poly (ethylene glycol) (PEG)-based hydrogel systems [51] has drawn attention. This newly developed hydrogel complex yields many benefits because of its conjugating activity, and it is difficult to induce bio-degradation into the monomers by natural metabolites, or cosmetic and medical substances. This biocompatible material was approved by the FDA in United States.
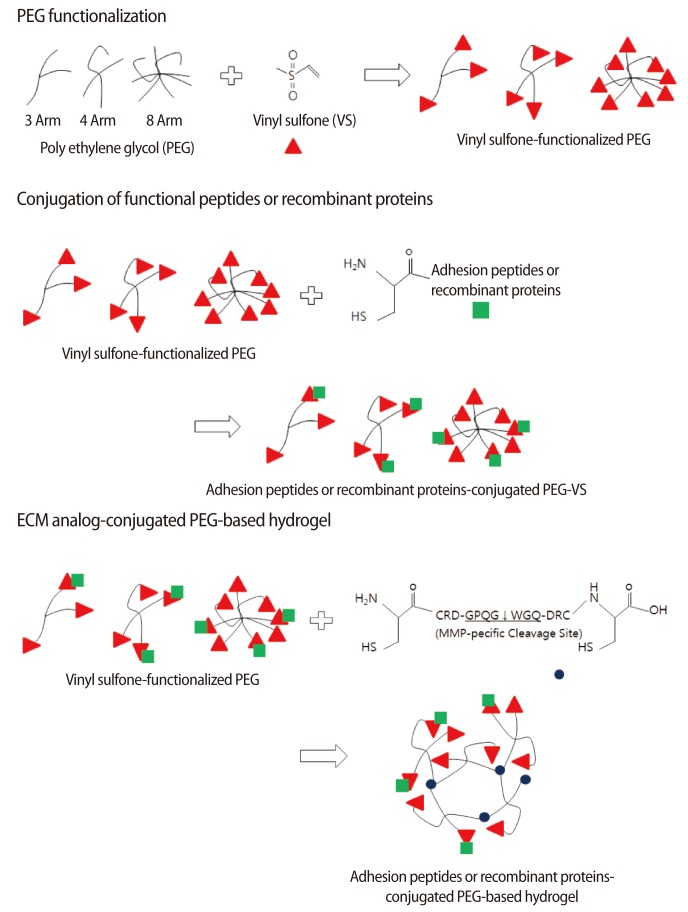
To create a cell-friendly microenvironment, a hydrogel polymerized by polyethylene glycol (PEG) has been suggested. In nature, the polymerization of diverse functional groups and functionalized PEG was conducted under cell-unfriendly conditions, which resulted in cellular toxicity. To avoid this insufficiency, a remodeled 3D scaffold system has been suggested (Figure 1). In this system, vinyl sulfone (VS)-functionalized PEG, which contains dicystein-containing peptide having matrix metalloproteinase (MMP)-specific cleavage sites (hereafter referred to as a combining region of the matrices with crosslinker), is employed. Therefore, conjugation of the sulfhydryl (SH) group of cystein with the VS group of PEG is conducted spontaneously through a Michael-type addition reaction in a cell-friendly environment [52]. Based on these previous achievements, employing VS-functionalized hydrogel is now being employed in the design of artificial niches for regulating stem-cell function and activity.
2. Designing 3D non-cellular niches with extracelular matrix analogs
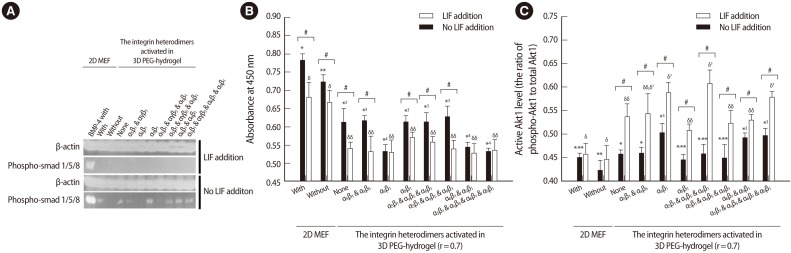
Using PEG-VS and a crosslinker, an ECM analog of an acellular niche has been developed and the microenvironmental influence of this 3D niche on self-renewal was evaluated in mouse ESCs (mESCs) [53]. The initial study evaluated the transcriptional, translational, and functional activity of integrin subunits and subsequently identified the expression of integrin ╬▒5╬▓1, ╬▒V╬▓5, ╬▒6╬▓1, and ╬▒9╬▓1 in the membranes of undifferentiated mESCs. These integrin heterodimers play important roles in ESC self-renewal and select RGDSP, TTSWSQ, and AEIDGIEL peptide motifs to co-activate integrin ╬▒5╬▓1, ╬▒V╬▓5, ╬▒6╬▓1, and ╬▒9╬▓1 (Figure 2). In next study, the effectiveness of PEG-based hydrogels of different mechanical properties, which were modulated by the ratio of (VS) and (SH) concentration (hereafter referred to as the stoichiometric ratio), was evaluated. The mechanical properties of the hydrogel with a 0.7 stoichiometric ratio significantly stimulated mESC self-renewal (Figure 3). Combinatorial conjugation of the optimized ECM analogs with selected motifs showed that four integrin heterodimers promoted self-renewal of mESCs. Subsequently, strong expression of stemness-related genes, which was similar to the expression under a conventional 2D cellular niche, was detected. Self-renewal of mESCs under the designed 3D acellular biomimetic niche was successfully maintained without stimulation of LIF signaling [54], and Stat3 activation by exogenous LIF is no longer the rate-limiting factor of stem cell self-renewal under a designated niche. Instead, Akt1/Smad signaling was significantly activated in the mESCs maintained under the 3D microenvironment (Figure 4). These data clearly indicate that a different pathway of ESC self-renewal is activated to adapt to a different microenvironment. Although significant changes in morphological properties were detected, this signal shift under a different niche does not cause any initial change in ESC self-renewal activity.
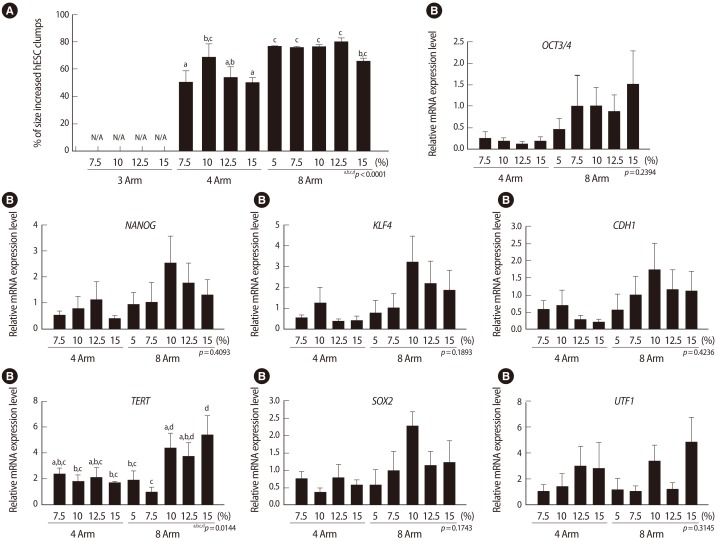
In the case of hESCs, a 3D acellular niche showed the potential to support hESC self-renewal without ECM analogs [54]. The optimal properties of 3D PEG-based hydrogel for the H9 hESC line was examined and the mechanical properties of the hydrogel were modified by the VS-functionalized PEG multiarm number and PEG concentration. It was found that hESC self-renewal and stemness were the highest after encapsulation in 8-multiarm PEG hydrogel consisting of 10% (wt/v) PEG (Figure 5). This optimized PEG-hydrogel was successfully applied to the 3D culture of H1 and Novo hESC lines. The 3D scaffold reduced the difference in the proliferative activity among three hESC lines evaluated. However, it does not seem that this optimized condition is sufficient for long-term maintenance of hESCs.
Conclusion
The possibility of maintaining stem cell self-renewal was found under a defined 3D niche, which is different from a conventional 2D niche. The PEG-based hydrogel using PEG-VS and crosslinkers containing both SH group and MMP-specific cleavage sites greatly contributes to establishing a cell-friendly 3D non-cellular niche system, which is capable of constructing diverse niches by mimicking different biomechanics and patterning after diverse ECM analogs. Probably, the suggested, hydrogel-nased system can be useful for creating cell-friendly environment in vitro. However, further trials to improve this 3D non-cellular niche for ESC self-renewal should be conducted for field and clinical application. There have been only a few applications using any 3D culture system, but more extensive studies about the characterization of stem cell niches in vivo. Attempts to synthesize ECM analogs or recombinant proteins, or to discover conjugating peptides will suggest a variety of alternative technologies for stem cell manipulation.