Introduction
Successful implantation depends on communication between the trophoblast of the blastocyst and the epithelial cells of the uterine endometrium. This communication is regulated by several inflammatory cytokines and adhesion molecules. Collectively, the process of implantation poses an immune challenge between the embryo (as a semi-allogenic) body and the uterus [1-3]. The maternal immune system promotes immune tolerance toward the embryo to maintain a normal pregnancy while defending against infection. The maternal immune response involves maintaining uterine receptivity, mediated by endometrial epithelial attachment molecules, trophoblast invasion, and extracellular matrix (ECM) breakdown [4,5].
Hormones, growth factors, and cytokines are crucial for the maintenance of uterine receptivity and implantation stages. Various cytokines, such as interleukin, leukemia inhibitory factor, and transforming growth factor-beta, are sensitive to local and systemic changes. They need to be appropriately regulated for successful implantation [6,7]. These cytokines are mostly mediated by the toll-like receptor (TLR) family, which is the main regulator of the immune response. Human endometrial tissue and trophoblasts express the TLR family [8]. The 10 membrane-spanning members of the TLR family play a critical role in modulating the inflammatory responses in humans. Each receptor responds to a specific ligand. Lipopolysaccharide (LPS) is a well-known endotoxin that consists of a lipid and a polysaccharide composed of O-antigen, outer core, and inner core joined by a covalent bond. LPS is a component of the cell wall of Gram-negative bacteria (e.g., Escherichia coli, Ureaplasma urealyticum, and Gardnerella vaginalis) [9,10]. LPS is an antigen that induces immune responses in uterine endometrial cells by TLR4 activation. LPS can trigger an imbalance in cytokines in the uterine endometrium [11,12].
Endometrial epithelial cells secrete hormones (estrogen and progesterone), growth factors, and various types of cytokines to enable successful implantation [13]. Adhesion molecules secreted by endometrial epithelial cells are an important factor in the embryo attachment stage [6,14]. Interactions between cells and the ECM are mediated by adhesion molecules [15], which consist of four subtypes: immunoglobulins, cadherins, integrins (ITGs), and selectins. ITGs are a major class of receptors within the ECM that mediate cellŌĆōECM interactions with collagen, fibrinogen, fibronectin, and vitronectin. ITGs provide essential links between the extracellular environment and intracellular signaling pathways, and are transmembrane receptors that mediate cell adhesion [16,17]. ITGs consist of two subunits: alpha (╬▒) and beta (╬▓). In humans, ITGs have 18 ╬▒ subunits and 8 ╬▓ subunits [18]. In this study, we analyzed ITG╬▒V, ITG╬▓3, and ITG╬▓5 to understand the effect of LPS on the attachment of trophoblastic spheroids to uterine endometrial epithelial cells.
Many in vitro models have provided insights into the implantation process [19,20]. In a recent study, in vitro implantation models were designed using the spheroid form of trophoblastic cells to surrogate embryos [21]. Trophoblast cell lines, including JAr, JEG-3, and human endometrial epithelial cells of ECC-1 were used as in vitro implantation models in previous studies [22-24]. This study was performed to investigate the detrimental effects of LPS on the attachment and outgrowth of various types of trophoblastic spheroids and endometrial epithelial cells as an in vitro model of implantation.
Methods
1. Culture of human trophoblastic and endometrial epithelial cells
The human trophoblastic JEG-3 cell line was cultured in DMEM (Welgene, Gyeongsan, Korea) and supplemented with 10% fetal bovine serum (FBS; Gibco, Waltham, MA, USA) and 1% penicillin-streptomycin (P/S; Lonza, Morristown, NJ, USA). The human trophoblastic JAr cell line and human endometrial epithelial cells (ECC-1 cells) were cultured in RPMI 1640 (Welgene) with 10% FBS and 1% P/S. Cells were cultured under standard conditions (37┬░C, 5% CO2), and the culture medium was replaced with fresh medium every 48 hours. LPS from E. coli O111: B4 (Sigma-Aldrich, St. Louis, MO, USA) was used to treat the ECC-1 cells to mimic endometrial infections caused by Gram-negative bacteria.
2. Preparation of various trophoblastic spheroids using the hanging drop method and the Organoid 3D culture kit
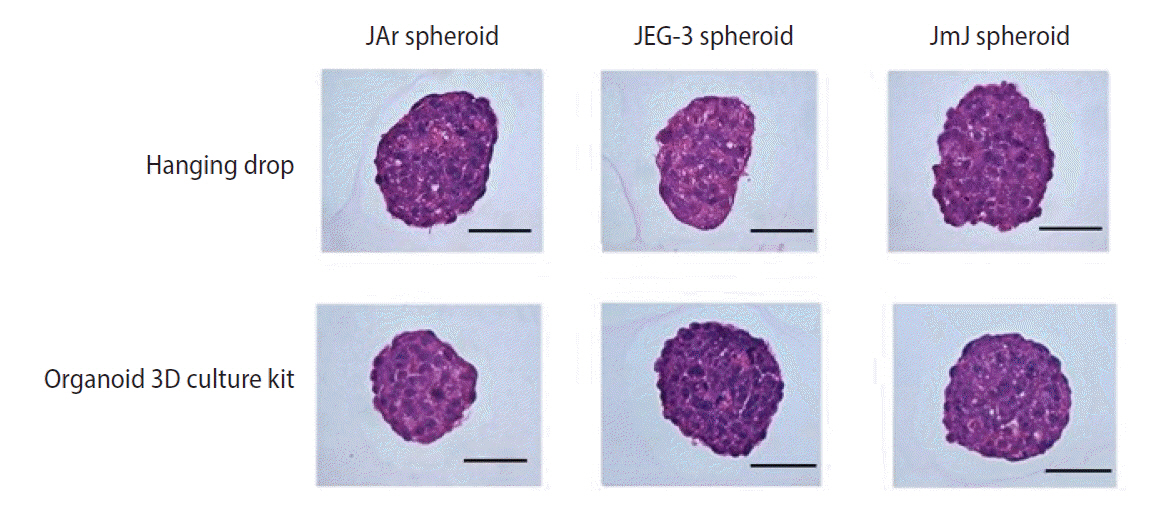
Spheroids of JAr, JEG-3, and JAr mixed JEG-3 (JmJ) were prepared using the hanging drop method and the Organoid 3D culture kit (Cell Smith, Seoul, Korea). JAr and JEG-3 cells were mixed at a 1:1 ratio to prepare JmJ spheroids. Using the hanging drop method, trophoblast cells (2 ├Ś104/20 ╬╝L drop) were plated onto the lid of a 100-mm dish in a regular array (20 drops/lid). The lid was inverted over the bottom, which was filled with Dulbecco's phosphate-buffered saline (DPBS; Biowest, Riverside, MO, USA), and cultured under standard conditions for 72 hours. Using the Organoid 3D culture kit, cells were seeded at a density of 1 ├Ś105 cells in 3 mL of the medium. The cells were incubated for 72 hours in the Organoid 3D culture kit. Spheroids sized between 200 and 300 ╬╝m were selected and used for subsequent experiments. The morphology of spheroids cultured by the different methods was observed using an EVOS XL Core Cell Imaging System (Thermo-Fisher, Waltham, MA, USA).
3. Histology of various types of trophoblastic spheroids
Three types of trophoblastic spheroids were fixed with 4% paraformaldehyde (Biosesang, Seongnam, Korea), and over 50 spheroids were suspended in 20 ┬ĄL of 2% agarose gel solution. The pre-embedded agarose gel blocks were embedded in paraffin, and sections of paraffin blocks with a thickness of 6 ┬Ąm were cut. The sections were placed on glass slides and stained with hematoxylin and eosin (H&E). The spheroids were observed under a Nikon Eclipse 80i microscope.
4. Quantitative analysis of mRNA expression in ECC-1 cells treated with LPS
Total RNA from the ECC-1 endometrial epithelial cells was isolated using TRIzol reagent (Ambion, Austin, TX, USA). Complementary DNA was synthesized using a cDNA reverse transcription kit (Takara Bio, Shiga, Japan). All quantitative SYBR-based quantitative real-time polymerase chain reaction (qRT-PCR) assays were performed in a 20 ╬╝L reaction volume using the StepOne software ver. 2.3 using the SYBR Green master mix, SensiFAST SYBR Hi-ROX Kit (Bioline, London, England), and 10 pM of each specific primer and 1 ╬╝L of cDNA per reaction (Table 1). Each qRT-PCR involved an initial denaturation step at 95┬░C for 10 minutes, followed by 40 cycles of denaturation at 95┬░C for 15 seconds, annealing at a specific temperature of each primer for 30 seconds, and extension at 72┬░C for 30 seconds. Finally, quantitative analysis was performed using the 2ŌĆō╬ö╬öct method with ╬▓-actin as an internal control [25].
5. Western blot analysis of ECC-1 cells treated with LPS
Equal amounts of total protein (20 ╬╝g) from ECC-1 cells treated with LPS were separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred to nitrocellulose membranes. The membranes were blocked with 5% skim-milk/TBS-T (Tris-buffered saline-Tween 20) solution (Bio-Rad, Contra Costa, CA, USA) and incubated with anti-ITG╬▒V (ab179475), ITG╬▓3 (ab197662), ITG╬▓5 (ab31327; Abcam, Cambridge, England), and anti-GAPDH (SC-32233; Santa Cruz Biotechnology, Dallas, TX, USA) antibodies. After the reaction with appropriate secondary antibodies linked to horseradish peroxide (Abcam), the signals were visualized using the ChemiDoc MP Imaging System (Bio-Rad). Densitometry was performed using ImageJ software (National Institutes of Health, Bethesda, MD, USA). Relative optical density was calculated by dividing the optical density of ITG protein by that of the internal control (GAPDH).
6. Evaluation of the attachment rate and outgrowth of trophoblastic spheroids on ECC-1
Endometrial epithelial cells of ECC-1 were cultured until confluence in T75 flasks and then detached using 0.25% trypsin/EDTA. Approximately 1 ├Ś105 endometrial cells were seeded in 12-well plate culture dishes and cultured at 37┬░C until 100% confluency. After LPS treatment in ECC-1 (0, 1, and 10 ╬╝g/mL of LPS, incubated for 24 hours), trophoblastic spheroids were added to ECC-1 and co-cultured for 6 hours. The attachment rate of trophoblastic spheroids to endometrial epithelial cells was evaluated at 0, 1, 2, 4, and 6 hours. The number of attached spheroids was counted by tapping by hand 3ŌĆō5 times. After 72 hours of co-culture, the outgrowth area and spheroid area were analyzed using ImageJ software (National Institutes of Health). The areas of outgrowth of the spheroids were measured in pixel units. The ratio of the outgrowth area (outgrowth area/spheroid area) was calculated and analyzed.
7. Statistical analysis
All experiments were performed at least in triplicate, and more than 250 spheroids were used in each group. Data are presented as mean┬▒standard error of the mean. The statistical significance of differences between groups was analyzed using one-way analysis of variance and the Tukey test. Data were analyzed using Prism GraphPad software ver. 5.0 (GraphPad, San Diego, CA, USA). Statistical significance was set at p<0.05.
Results
1. Morphology and histology of trophoblastic spheroids using the hanging drop method and Organoid 3D culture kit
Microscopic analysis of the various types of spheroids was performed to obtain more detailed observations of their morphology. Spheroid formation and cell distribution were observed histologically by H&E staining. The spheroids cultured by both methods (hanging drop and the Organoid 3D culture kit) were well prepared for the in vitro implantation model. All trophoblastic spheroids were maintained at a high cell density (Figures 1 and 2).
2. Effect of LPS on mRNA expression of adhesion molecules and inflammatory cytokines in ECC-1 cells
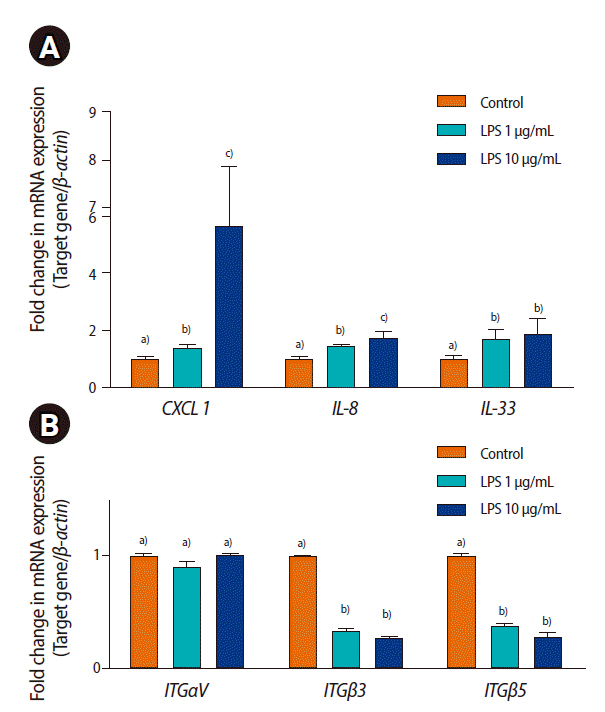
To evaluate the effect of LPS on ECC-1 cells, the expression of inflammatory cytokines and adhesion molecules was analyzed by qRT-PCR. LPS treatment significantly increased the mRNA expression of CXCL1, IL-8, and IL-33 and decreased ITG╬▓3 and ITG╬▓5 expression in ECC-1 cells (p<0.05). However, the mRNA expression of ITG╬▒V in endometrial epithelial cells was not changed by LPS treatment (Figure 3).
3. Effect of LPS on protein expression of adhesion molecules in ECC-1
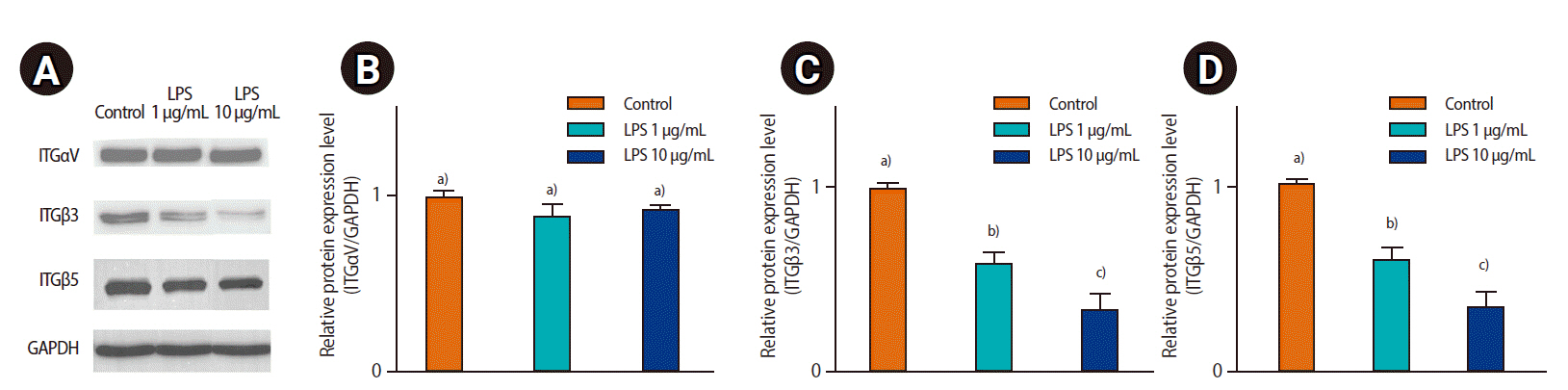
To evaluate the effect of LPS on ECC-1, the expression of adhesion molecules (ITG╬▒V, ITG╬▓3, and ITG╬▓5) was analyzed by western blotting. ITG╬▓3 and ITG╬▓5 expression decreased in response to LPS treatment. However, ITG╬▒V expression was not significantly different between the control and LPS-treated groups (Figure 4).
4. Effect of LPS on the attachment of various trophoblastic spheroids on ECC-1
The attachment rate between trophoblastic spheroids and the ECC-1 endometrial epithelial cells was evaluated in a time-dependent manner. The attachment rate was evaluated under 1% FBS or without FBS. LPS treatment did not affect the attachment rate of JEG-3 spheroids (Table 2). However, the attachment rates of JAr spheroids to ECC-1 cells were significantly decreased by LPS treatment (p<0.05). The attachment rates of JAr spheroids on ECC-1 were similar in the LPS-treated and control groups after 6 hours of co-culture (Table 3). In the absence of FBS supplementation, the attachment rate was significantly decreased by LPS treatment compared to the condition with 1% FBS supplementation (p<0.05). In the following experiments, the attachment rate of JmJ spheroids was analyzed in the absence of FBS supplementation. In JmJ spheroids, the attachment rate of LPS treatment was significantly lower than that of the control after 2 hours of co-culture, as shown in Table 4 (p<0.05).
5. Effect of LPS on the outgrowth of various trophoblastic spheroids on ECC-1 cells
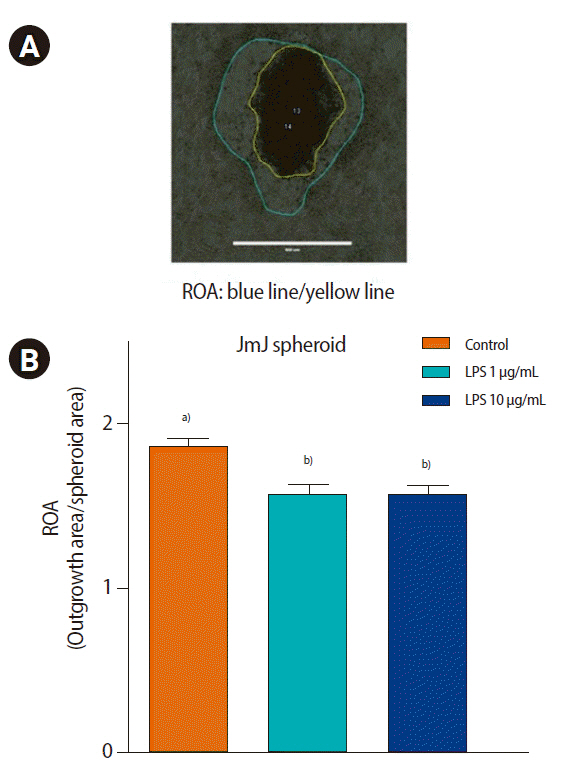
The outgrowth of trophoblastic spheroids on the ECC-1 endometrial epithelial cells was evaluated after 72 hours of co-culture. The JAr spheroids did not show any outgrowth area, whereas the JEG-3 and JmJ spheroids showed outgrowth areas on ECC-1. Nevertheless, LPS treatment did not affect the outgrowth area of JEG-3 spheroids. Interestingly, LPS treatment significantly decreased the outgrowth area of JmJ on ECC-1 cells after 72 hours of co-culture, as shown in Figure 5 (control, 1.90┬▒0.06; 1 ╬╝g/mL LPS, 1.62┬▒0.06; and 10 ╬╝g/mL LPS, 1.64┬▒0.06; p<0.05).
Discussion
Implantation is a complex immunological process. The inflammatory environment of the uterus changes from a pro-inflammatory to an anti-inflammatory state throughout implantation and pregnancy. A strong inflammatory response is necessary during implantation [26-30]. However, an imbalance in the immune status of the uterus can cause serious problems such as implantation failure and pregnancy loss [31,32]. In this study, activation of TLR4 in ECC-1) by LPS significantly reduced the attachment rate and outgrowth area between trophoblastic spheroids and endometrial epithelial cells.
It has been shown that implantation failure is induced by LPS through stimulation of the innate immune system and activation of the TLR4 pathway in the early stages of pregnancy [11,33]. A similar study suggested that TLR3 stimulated by a synthetic ligand, poly I: C (which is a double-stranded RNA molecule) in CBA/J female mice increased fetal losses [34] and reduced actin polymerization and adhesion molecule expression in endometrial cells [35]. In another study, activation of TLR5 in a human telomerase immortalized endometrial epithelial cell line (hTERT-EECs) by bacterial flagellin significantly decreased the attachment rate between JAr spheroids and underlying endometrial cells [36]. This study suggested that activation of the TLR family by various antigens at early stages of pregnancy could induce detrimental effects on implantation.
An endometrial epithelial cell line, ECC-1, was treated with LPS to mimic endometrial infections caused by Gram-negative bacteria. In the implantation process, LPS affects not only endometrial epithelial cells, but also competent blastocysts [37]. LPS stimulates the TLR4 pathway to activate the inflammatory response [38]. TLR4 is essential for LPS activation. However, the JAr and JEG-3 human trophoblast cell lines do not express TLR4 on the membrane [39]. Other regulated pathways could potentially affect trophoblasts, resulting in the detrimental effect of LPS. Human endometrial epithelial cell lines (Ishikawa and ECC-1 cells) expressing TLR4 were used to evaluate the attachment rate of trophoblastic spheroids by LPS treatment [40]. In a pilot study, endometrial epithelial cell lines (Ishikawa and ECC-1 cells) were used in the implantation model to investigate the effects of LPS. LPS treatment did not affect the attachment rate between Ishikawa cells and trophoblastic spheroids. However, the attachment rate between ECC-1 cells and trophoblastic spheroids of JAr decreased with LPS treatment. Thus, we used EEC-1 endometrial epithelial cells in subsequent experiments of the implantation model.
The attachment rate of JAr and JEG-3 spheroids with ECC-1 cells was evaluated in the presence of 1% FBS and without FBS supplementation. In our experiments, the detrimental effect of LPS could be more clearly observed in the absence of FBS supplementation than in the 1% FBS condition due to the fact that FBS contains various factors such as growth factors and adhesion molecules [41]. Bas et al. [42] showed that FBS contains unknown factors and can inhibit TLR activation. For this reason, we designed the attachment and outgrowth experiments of JmJ spheroids without FBS supplementation.
The spheroids prepared by both methods (hanging drop and the Organoid 3D culture kit) were observed histologically using H&E staining. The microscopic observations showed that the Organoid 3D culture kit could provide spheroids with more consistent size and circularity than the hanging drop method. Therefore, spheroids prepared with the Organoid 3D culture kit might be more reliable for growth assays that analyze the ratio of the growth area to the spheroid area. We attempted to identify distinct features of JmJ spheroids compared to JAr and JEG-3 spheroids by histological observations. However, no significant differences were observed between the groups.
Choriocarcinoma JAr and JEG-3 cell lines were used to establish an in vitro model to investigate the attachment of trophoblast cells. JAr spheroids were suitable for investigating the effect of LPS on the attachment rate. However, the JAr spheroids did not show any outgrowth areas, indicating that they were not suitable for outgrowth assays. The other trophoblastic spheroids, JEG-3, had a clear outgrowth area for outgrowth assays. However, LPS-treated ECC-1 cells showed no significant difference in the attachment rates of JEG-3 spheroids compared to the control group.
Notably, JmJ spheroids were suitable for both attachment rate and outgrowth assays with ECC-1 cells. The JmJ spheroids showed a significant difference in the attachment rate after treatment with LPS. Moreover, the JmJ spheroids had a clear outgrowth area, which was suitable for outgrowth assays. The JmJ spheroids were used for subsequent experiments involving LPS treatment.
In the attachment rate assay, all spheroids were attached to ECC-1 after 6 hours of co-culture. Usually, the trophoblastic cells of carcinoma cell lines are used for in vitro implantation models. Carcinoma cell lines have a stronger invasion ability than normal cell lines [43]. Taken together, using a highly invasive carcinoma cell line could mask the detrimental effects of LPS on the attachment rate after 6 hours of co-culture of trophoblastic spheroids and ECC-1.
The expression of inflammatory cytokines and adhesion molecules was analyzed in the present study. ITGs are adhesion molecules in endometrial, decidual, and extravillous cytotrophoblast cells. They participate in cell-cell adhesion and adhesion between cells and components of the ECM [44]. LPS treatment significantly decreased the mRNA expression of ITG╬▓3 and ITG╬▓5. However, the mRNA levels of ITG╬▓1 and ITG╬▒V were not changed by LPS treatment. A recent study by Guo et al. [45] using a bovine model could help understand these results. The expression of ITG╬▓3, ITG╬▓5, ITG╬▓7, and most transcripts coding for cell adhesion molecules (PCDH7, PKP1, PKP3, CT, CTNNA3, CTNNAL1, and CDH2) were downregulated after treatment with 2 ╬╝g/mL LPS in bovine endometrial epithelial cells. However, transmembrane glycoproteins that mediate cell-cell interactions through calcium binding (i.e., ITGB6, CDH26, ITGAV, and CELSR1) were overexpressed after LPS treatment [46]. In a preliminary study, we evaluated the relative expression of ITG╬▓3 and ITG╬▓5 mRNA to determine whether LPS treatment might decrease the attachment between spheroids and ECC-1 (data not shown). However, there were no significant differences among the different types of spheroids (JAr, JEG-3, and JmJ).
In this study, the mRNA expression of the inflammatory cytokines CXCL1, IL-8, and IL-33 was higher in LPS-treated ECC-1 cells. CXCL1 and IL-8 are representative inflammatory cytokines in endometrial cells exposed to LPS. IL-8 and CXCL1 are cytokines with neutrophil chemotactic and activating activity and T cell chemotactic activity that play a major role in the recruitment of leukocytes to the endometrium via the CXCR1 and CXCR2 pathways [46-48]. However, the relationship between IL-33 and LPS in endometrial epithelial cells remains unclear. In a study by Miller et al. [49], IL-33 stimulated the expression of inflammatory cytokines, such as CXCL1, IL-6, and IL-15, in the progression of endometriosis. Further studies are needed to understand the relationship between the inflammatory response in endometrial cells and IL-33 expression.
Our study had some limitations. Numerous factors are involved in the implantation process in vivo, including immune cells, stromal cells, and hormones [50]. However, in this study, only trophoblastic cells and endometrial epithelial cells were used because of the limitations of the in vitro model. Including various other factors in the in vitro model would help to understand the details of the implantation process.
This study had several advantages. The implantation process is highly complex and difficult to mimic in vitro. Many researchers have used embryo transfer to assess implantation potential. However, embryo transfer into pseudopregnant mice requires expert experimental skills and numerous sacrificial animals [51-53]. An in vitro implantation model using the novel trophoblastic JmJ spheroids and ECC-1 endometrial epithelial cells could overcome these problems.
The findings of our study could help to understand the detrimental effects of LPS on the attachment and outgrowth of various types of trophoblastic spheroids and endometrial epithelial cells in an in vitro model of implantation (Figure 6). The results of this study suggest that alterations in the expression levels of inflammatory cytokines (CXCL1, IL-8, and IL-33) and adhesion molecules (ITG╬▓3 and ITG╬▓5) by LPS treatment might be related to reduced trophoblastic spheroid attachment and outgrowth on endometrial epithelial cells.