Introduction
Endometriosis, defined as the presence of endometrial glandular and stromal cells in sites outside the uterine cavity, is an estrogen-dependent inflammatory disease. It affects 5% to 10% of women of reproductive age, and the main clinical features are chronic pelvic pain, dyspareunia, and infertility [1]. Although the pathogenesis of endometriosis has not been elucidated, endometriosis is a highly proliferative and benign gynecologic disorder with the ability to undergo neoplastic-like processes. Endometriotic lesions have aberrant overexpression of anti-apoptotic genes, such as bcl-2 [2], various cellular adhesion molecules, and metalloproteinases [3] and low expression of pro-apoptotic genes, such as bax [4]. In addition, molecular distinctions, such as the overproduction of estrogen, prostaglandins [5], and cytokines [6], also occur in the endometrium of women with endometriosis relative to the endometrium of women without the disease. These conditions are able to enhance the adhesion of shed endometrial tissue onto the ectopic site and the progression of disease.
Endometriosis shares a predisposing factor and molecular similarity with cancer [7]. Increased telomerase activity has been detected in various cancers, such as endometrial and ovarian cancer [8,9], and is also related to a sustained replication potential in endometriosis [10]. Telomeres are the non-coding tandemly repeated DNA sequences (5-TTAGGG-3) that protect all eukaryotic chromosomal ends. They are markers for cellular aging, as their length declines with each cell division. Most human cancer cells maintain their telomeres through activation of telomerase. Telomerase is a ribonucleoprotein complex and plays an important role in telomere maintenance and cellular immortality, and its core functional component is human telomerase reverse transcriptase (hTERT). A correlation between hTERT mRNA expression and telomerase activity has been observed [11]. An immunohistochemical study has suggested that aberrant endometrial expression of telomerase enhances endometrial cell proliferation, contributing to the pathogenesis of endometriosis [12].
Peroxisome proliferator activated receptor γ (PPARγ) is a pleiotropic nuclear hormone receptor that binds to specific DNA response elements and may regulate gene expression indirectly, negatively, or positively, through competition with other transcription factors [13]. Using a DNA array containing 96 genes known to control the cell cycle, PPARγ inhibited expression of genes encoding the minichromosome maintenance proteins (MCM) 6 and 7 [14]. MCM proteins play a central role in the regulation of the initiation of DNA replication and ensure that DNA replicates only once during each cell cycle [15]. Therefore, PPARγ itself is considered to be a novel important regulator of the cell cycle and apoptosis and may provide a new therapeutic approach to the treatment of proliferative cardiovascular diseases and cancer [16-18]. Several reports have shown that PPARγ agonist (rosiglitazone) treatment induced not only growth inhibition of endometrial explants [19] but also regression of established lesions [20]. We previously reported that rosiglitazone inhibited the phosphorylation of Smad and extracellular signal-regulated kinase (ERK) in human endometrial stromal cell (ESC) decidualization that is induced by TGF-β1 in vitro [21]. During this process, rosiglitazone was able to inhibit PGE2 production through the down-regulation of the ERK pathway. Local positive feedback loops of estrogen and cytokines are essential for ESC proliferation in eutopic endometrium with endometriosis.
Based on the above considerations, we have examined the effects of PPAR γ activation by rosiglitazone on the expression of hTERT, a core functional component of telomerase activity, and on the local cytokine production, prostaglandin E2 (PGE2) pathway through ERK phosphorylation in in vitro cultured eutopic ESCs from patients with endometriosis.
Methods
1. Clinical subjects and endometrial biopsies
Human endometrial tissues were obtained from patients with endometriosis, which was confirmed by laparoscopic operation, and without endometriosis (control group) via endometrial biopsy. Biopsies were performed during operative laparoscopy at Ajou University Hospital from April 2007 to June 2008. Informed consent was obtained from each participating patient, and the study was approved by the Institutional Review Board (AJIRB-GEN-SMP-10-187). Tissue specimens were acquired from patients aged 30 to 45 who were estimated to be in the mid or late proliferative phase of the menstrual cycle. Five subjects were enrolled in each group, and patients with endometriosis stage III-IV were included in the study group.
2. Reagents and antibodies
The PPAR γ agonist, rosiglitazone, was purchased from Alexin (Alexin, San Diego, CA, USA). The phosphorylated ERK (pERK) inhibitor PD98059 was purchased from Calbiochem (Calbiochem, San Diego, CA, USA). The antibodies against human pERK/ERK (mouse), hTERT (rabbit), COX-1, 2 (goat), and aromatase (goat) were purchased from Santa Cruz Biotechnology (Santa Cruz Biotechnology, Santa Cruz, CA, USA). The antibody against b-actin was purchased from Sigma. The antibodies against goat horseradish peroxidase (HRP), mouse HRP, and rabbit HRP were purchased from Chemicon (Chemicon, Temecula, CA, USA).
3. Endometrial stromal cell isolation and primary cell culture
The protocol for cell isolation was identical to that of a previously reported method [22]. Briefly, the endometrial tissues obtained were placed in a conical tube (Falcon, Becton Dickinson, Franklin Lakes, NJ, USA) containing Dulbecco's modified Eagle's medium (DMEM)/F-12 supplemented with 10% FBS (Gibco Life Technologies, Grand Island, NY, USA) and transported to the aseptic laboratory, where the tissue was again washed several times with phosphate-buffered saline (PBS) to remove any residual blood clots and FBS. Then the tissues were minced into small pieces, incubated in 10 mL of 0.5% trypsin-EDTA (Gibco Life Technologies) for 1 hour at 37℃. After incubation, 1 mL of FBS was added to stop the enzymatic reaction. The tissues were centrifuged at 1,200 rpm for 5 minutes. The pellets were suspended with 5 mL of DMEM/F-12 and centrifuged at 1,200 rpm for 5 minutes. The pellets were resuspended in 10 mL of DMEM/F-12 containing 10% FBS, plated onto 100 mm dishes, and then allowed to adhere at 37℃ for 1 hour. Non-adhering epithelial cells and blood cells were removed by rinsing the cells with PBS. The adhering ESC were cultured in DMED/F12 containing 10% FBS, penicillin, and streptomycin. When the cells became confluent, they were dissociated with 0.5% trypsin-EDTA, harvested by centrifugation at 1,200 rpm for 5 minutes, seeded in 100 mm dishes at 1×106 cells, and then incubated at 37℃ in humidified 5% CO2. After incubation for 24 hours, the media was replaced with fresh media and cultured for 72 hours under the following conditions: varying doses of rosiglitazone (PPARγ-specific ligand, 2-40 µM). The proliferation of ESCs and expression of aromatase were dose-dependently suppressed by rosiglitazone. We chose a 20 µM dose of rosiglitazone for the follow-up experiments; three groups used 20 µM of rosiglitazone, or 20 mM of the pERK inhibitor (PD98059), or 20 µM of rosiglitazone and 20 mM of the pERK inhibitor (PD98059) together. The doses of agents added to the cultures were based on the study of Han et al. [23]. To determine the purity of the stromal cell preparations, they were characterized using immunostaining with antibodies to cytokeratin for epithelial cells (Santa Cruz Biotechnology) and vimentin for stromal cells (Santa Cruz Biotechnology). The purity of the stromal cells was over 98%, as judged by positive cellular staining for vimentin and negative cellular staining for cytokeratin.
4. Cell proliferation assays (MTT assays)
Cells were plated in 24-well plates at 5×104 cells/well and cultured at 37℃ for 24 hours. The media was replaced with fresh media and cultured for 72 hours under the following conditions: varying doses of rosiglitazone (PPARγ-specific ligand, 2-40 µM), 20 µM of rosiglitazone, and 20 mM of the pERK inhibitor (PD98059), and 20 µM of rosiglitazone and 20 mM of the pERK inhibitor (PD98059) together. After 72 hours of treatment incubation, the cells were subjected to a cell proliferation assay by using the 3-[4,5-dimethylthiazol-2-yl]-2,5 diphenyl tetrazolium bromide (MTT) Cell Proliferation Assay Kit (R&D system, Minneapolis, MN, USA). To each well, MTT reagent (final concentration, 0.05%) was added, followed by an additional 4 hours culture. Thereafter the culture media were removed, and the formazan formed was dissolved in 150 µL of dimethyl sulfoxide (DMSO), vortexed, and spectrophotometrically measured at 590 nm. Three sets of each group were subjected to MTT assay and each assay was repeated at least three times. The proliferation index was calculated: experimental optical density (OD) value×100/control OD value.
5. Western blot analysis of hTERT, aromatase, COX-2, and pERK
Western blot analysis was performed as described [24]. Briefly, cell lysates were prepared by extracting protein with lysis buffer (40 mM Tris-HCL [pH 8.0], 120 mM NaCL, 0.1% Nonider P-40) supplemented with protease inhibitors. After sonication, the whole-cell lysate was collected by centrifugation at 14,000 rpm at 24℃ for 20 minutes. The total proteins were separated by sodium dodecyl sulphate-polyacrylamide gel electrophoresis and transferred onto a nitrocellulose blotting membrane (Sartorius, Goettingen, Germany). The membranes were blocked with 5% nonfat dry milk in Tris-buffered saline and incubated with primary antibodies for 1 hour at room temperature (RT). The blots were developed by peroxidase-conjugated secondary antibody, and the proteins were visualized by an enhanced chemiluminescence procedure (Santa Cruz Biotechnology) according to the manufacturer's recommendations. Chemiluminescence measurements and semi-quantitative values were obtained by an Intelligent Dark Box II (Fujifilm, Tokyo, Japan) and Image Gauge V3.12 software (Fujifilm, Tokyo,Japan). Quantification of the western blots was performed by densitometry and normalization to β-actin and constitutive COX-1. The densitometric analyses were repeated three times using Image Gauge V3.12 (Fujifilm, Tokyo,Japan).
6. Measurement of PGE2 concentrations in the conditioned media
To measure the production of PGE2 during culture in various conditions for 72 hours, the cells were removed by centrifugation, and the cell-free conditioned media were collected and stored at -75℃ until use. After thawing at room temperature, the medium was analyzed for immunoreactive PGE2 by using anti-PGE2 monoclonal antibodies (PGE2 EIA kit, Cayman Chemicals Ann Arbor, MI, USA), according to the manufacturer's specifications, with absorbance at 450 nm in a plate reader (SpectraMax 190, Molecular Device, Sunnyvale, CA, USA). All of the experiments were repeated at least three times with different media under the same conditions to minimize intra- and interassay variations. The intra- and interassay coefficients of variation were <3.8% and <7.8%, respectively, for PGE2.
7. Statistical analysis
All of the experiments were performed at least three to four times in duplicate using independent cell cultures. Where appropriate, the data are expressed as mean±SM and statistically analyzed with an unpaired Student's t-test and Kruskal-Wallis one-way analysis of variance with Duncan's test, using the computer software program SPSS ver. 11.0 for Windows (SPSS Inc., Chicago, IL, USA). p<0.05 was considered statistically significant.
Results
1. ESC proliferation and hTERT expression in endometriosis
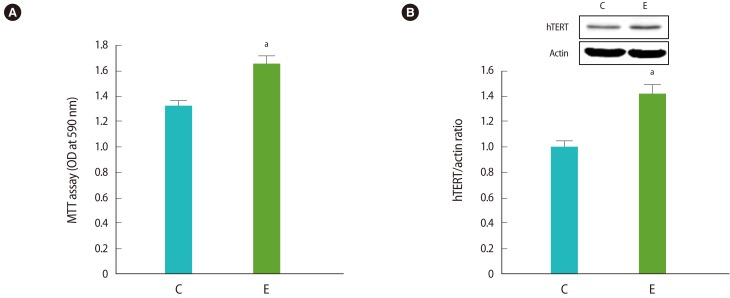
Cell proliferation was significantly increased in ESCs cultured in vitro from patients with endometriosis compared to those from controls (without endometriosis) by MTT assay. The protein expression of hTERT was also significantly increased in ESCs cultured in vitro from patients with endometriosis compared to those of the controls (Figure 1).
2. PPARγ agonist inhibits proliferation and hTERT expression in ESCs from endometriosis
To investigate the regulation of telomerase activity and cellular proliferation of eutopic endometrium by PPARγ agonist in endometriosis, cultured ESCs from patients with endometriosis were treated with different concentrations of a PPARγ agonist, rosiglitazone. As shown in Figure 2A, the PPARγ agonist also dose-dependently decreased the cell proliferation. Furthermore, isolated ESCs that were obtained from patients with endometriosis resulted in a substantial induction of hTERT expression. The hTERT expression was also significantly inhibited by the PPARγ agonist in a dose-dependent pattern (Figure 2B).
3. The effect of PPARγ agonist on the expression of aromatase, COX-2, and PGE2
To confirm the effect of PPARγ agonist on the basic pathologic processes of endometriosis, we focused on the expression of aromatase in the estrogen biosynthetic pathway and COX-2 expression in the prostaglandin pathway. During the culture of isolated ESCs from endometriosis, stromal cells were maintained under serum-free conditions for 24 hours and then treated with PPARγ agonist (20 µM) for 72 hours in the rosiglitazone group. The expression of aromatase was dose-dependently inhibited after the rosiglitazone treatment (Figure 2B). COX-2 is known to be a key enzyme of PGE2 production, and thus we also measured the concentration of PGE2 in the conditioned cultured medium using an enzyme linked immunosorbent assay (ELISA) assay. Although the inhibition of COX-2 expression by PPARγ agonist was not statistically significant, a subtle decrease in COX-2 could significantly suppress the PGE2 formation in ESCs with endometriosis (Figure 3). We attempted to explore the effect of rosiglitazone on the MAPK signaling pathway. This pathway includes two closely related MAPKs: extracellular-signal-related kinase 1 (ERK1) and 2 (ERK2). These two kinases are activated by phosphorylation; their activity was assessed by immunoblot analysis using antibodies against phosphorylated ERK1 and ERK2. As shown in Figure 3, the level of phosphorylated ERK1 and ERK2 expression was suppressed by rosiglitazone in eutopic ESCs of endometriosis.
4. Synergistic effect of PPARγ agonist and pERK inhibitor
To further determine whether the observed inhibition of COX-2 function (PGE2 production) in ESCs of endometriosis by PPARγ agonist is mediated through the ERK pathway, the biologically active form of phosphorylated ERK inhibitor (PD98059) was added to PPARγ agonist during the culture of isolated ESCs from endometriosis. Rosiglitazone treatment alone significantly inhibited cell proliferation and PGE2 production, coupled with suppression of pERK in ESCs from endometriosis (Figure 4).
As depicted in Figure 4A, the cell proliferation of ESCs was suppressed more by rosiglitazone alone than treatment with pERK inhibitor alone. Furthermore, co-treatment of rosiglitazone and pERK inhibitor inhibited ESC proliferation more than treatment with each agent alone. The concentration of PGE2 in the conditioned medium was also significantly decreased by co-treatment with rosiglitazone and pERK inhibitor (PD98059) compared with rosiglitazone or pERK inhibitor alone. This means that rosiglitazone and pERK inhibitor have a synergistic effect on the suppression of cell proliferation and PGE2 production in eutopic endometrium with endometriosis. In addition, both rosiglitazone and ERK inhibitor suppressed the expression of hTERT, aromatase, and COX-2.
Discussion
The purpose of this study was to investigate whether rosiglitazone, a PPARγ agonist, could impede the growth of ESCs from endometriosis during in vitro culture, and to elucidate the effect of PPARγ on eutopic endometrium of endometriosis. Endometriosis and ovarian cancer share common antecedent mechanisms or predisposing factors (e.g., genetic susceptibility, immune/ angiogenic dysregulation, environmental toxin exposure), with obvious divergence in molecular pathways downstream [7]. The eutopic endometrium of endometriosis has multiple aberrant molecular abnormalities, including the activation of oncogenic genes [25,26], overproduction of estrogen, cytokines, prostaglandins, and metalloproteinases [1], and abundant expression of angiogenic factors [27]. There have been several studies suggesting that the overexpression of fibroblast growth factors and anti-apoptotic factors such as Bcl-2 could contribute to favoring the survival and replenishment of endometriotic tissue outside of the uterus [26]. We have focused on the increased proliferation activity and previously reported an increased incidence of endometrial polyps in patients with endometriosis [28].
The replication potential and local positive feedback loop of estrogen in eutopic endometrium is essential for ESC proliferation. hTERT is a core functional component of telomerase activity. Increased telomerase expression correlates with cell proliferation [29] and most endometrial cancers [30]. It has been reported that over-expression of hTERT mRNA and telomerase activity in the endometrium of endometriosis patients and the levels of hTERT mRNA and telomerase activity increased as the disease became more severe [10]. It is suggested that the replication potential of regurgitated endometrial tissue may play an important role in the pathogenesis of endometriosis. A positive correlation has been shown between hTERT mRNA expression by quantitative real-time RT-PCR and telomerase activity by TRAP assay in endometrial tissue [10,11]. Therefore, we investigated the hTERT protein expression of cultured ESCs as an alternative to observing the telomerase activity in woman with endometriosis.
PGE2 was found to be the most potent known inducer of aromatase activity in ESCs [5], and estrogen increased PGE2 formation by stimulating COX-2 enzyme in ESCs in vitro [31]. Thus, a positive feedback loop for the continuous local production of estrogen and PGE2 has been established, leading to the proliferative and inflammatory activity of endometriosis. In the eutopic endometrium of women with endometriosis, there is a subtle increase in COX-2 activity and detectable aromatase activity. Therefore, moderate PGE2 and low estradiol can be detected locally. In ectopic endometriotic tissue, full-blown molecular abnormalities include high COX-2 and aromatase levels. Excessive formation of PGE2 in endometrial and endometriotic tissues can cause severe menstrual cramps and chronic pelvic pain. The estradiol level in endometrial tissue is high because estradiol is overproduced by aromatase and is not metabolized owing to deficient HSD17B2 activity [32]. Yoshino et al. [33] reported that ERK is present in endometriotic tissues and is activated by pro-inflammatory agents. They also showed that IL-1-induced COX-2 expression was suppressed by ERK inhibitors. This finding suggests that the ERK pathway may play a possible pathophysiologic role in endometriosis, acting as an intracellular signal transducer.
PPARγ agonists have been shown to mediate the pro-apoptotic activity of vascular smooth muscle cells [34], and inhibit telomerase activity during the proliferative response after vascular injury [16], and have thus provided a new therapeutic approach to preventing restenosis. Therefore, we attempted to elucidate the effect of PPARγ agonist on hTERT expression, and the mechanisms underlying the interaction of PPARγ agonist and production of the local cytokine and PGE2 pathway through ERK phosphorylation in the eutopic endometrium of endometriosis. We observed that PPARγ agonist dose-dependently decreased the cell proliferation and hTERT expression. The expression of aromatase was also dose-dependently inhibited after rosiglitazone treatment. Our results agree with the finding that PPARγ agonist decreases aromatase activity in cultured human granulosa cells [35]. In addition, PPARγ agonist decreased phosphorylated ERK in the eutopic endometrium of endometriosis. The release of PGE2 by cultured ESCs was significantly decreased. The expression of aromatase and COX-2 was significantly inhibited after the addition of rosiglitazone, and this resulted in a considerable decrease in PGE2 production in the culture medium. These changes were decreased more by adding ERK inhibitor. This means that rosiglitazone and pERK inhibitor have a synergistic effect on the suppression of cell proliferation and PGE2 production in eutopic endometrium with endometriosis. Similarly, we have also noted that PPARγ agonist is linked to a negative role in ERK signaling, which is activated by TGF-β in the decidualization process of cultured normal human ESCs [21].
In fact, few animal studies concerning PPARγ agonist were able to find a regression of endometriotic lesions. It has been shown that a PPARγ agonist, rosiglitazone, with the ability to modulate inflammatory cells and cytokine formation, affected the induction of endometriosis negatively in an experimental rat model [19], and the regression of experimental endometriosis in rat [20,36] and baboon models [37]. PPARγ ligands reduced monocyte migration in an in vitro study of the peritoneal fluid of women with endometriosis [38]. Ciglitazone has been shown to decrease the number of peritoneal macrophages in a mouse model [39] and to decrease attachment of endometrial cells to peritoneal mesothelial cells in an in vitro model of early endometriotic lesions [40]. In our study, we found that PPARγ agonist could diminish PGE2 production from eutopic endometrium, which was thought to be the cellular origin of endometriosis. This could be explained by a regression of the endometriotic lesion in an animal model by a PPARγ agonist, which resulted in the decrease in hTERT, aromatase, and COX-2 expression. In addition, several studies have shown that PPARγ agonists could decrease aromatase activity in cultured human granulosa cells [35] and immunomodulate endometrial epithelial and stromal cells [38,41]. PPARγ agonists in animal models of endometriosis have shown promise; further studies could be undertaken in efforts to determine the clinical efficacy of PPARγ agonists as a potential treatment option for endometriosis.
The safety of PPARγ agonists in early pregnancy has been suggested by recent studies. Studies in mice have shown that rosiglitazone does not have an adverse impact on embryo or neonatal development [42], and may even have beneficial effects on blastocyst quality in female mice with diet-induced obesity [43]. A small case series of pregnant women with polycystic ovary syndrome who took rosiglitazone during the first trimester of pregnancy demonstrated no evidence of developmental or congenital abnormalities [44]. It should be mentioned that the meta-analysis suggesting that the PPARγ agonist rosiglitazone increases the risk of cardiac ischemia among type 2 diabetics [45] shifted the potential extension of the results of our study to the clinical use of pioglitazone, a PPARγ agonist that has been shown to have a favorable safety profile in type 2 diabetics, in terms of cardiovascular health [46].
Overall, our study suggests that the PPARγ agonist rosiglitazone suppressed cell proliferation and hTERT expression in in vitro cultures of eutopic ESCs of endometriosis. Moreover, the expression of aromatase and COX-2 was significantly inhibited after the rosiglitazone treatment, resulting in a considerable decrease in PGE2 production in culture medium through the partial inhibition of the ERK pathway. Further clinical studies in humans could be conducted on whether these results could be translated into reduced pelvic pain or endometriotic lesion size, such as endometrioma, with rosiglitazone treatment.