Glucose-6-phosphate dehydrogenase deficiency does not increase the susceptibility of sperm to oxidative stress induced by H2O2
Article information
Abstract
Objective
Glucose-6-phosphate dehydrogenase (G6PD) deficiency is the most common human enzyme defect. G6PD plays a key role in the pentose phosphate pathway, which is a major source of nicotinamide adenine dinucleotide phosphate (NADPH). NADPH provides the reducing equivalents for oxidation-reduction reductions involved in protecting against the toxicity of reactive oxygen species such as H2O2. We hypothesized that G6PD deficiency may reduce the amount of NADPH in sperms, thereby inhibiting the detoxification of H2O2, which could potentially affect their motility and viability, resulting in an increased susceptibility to infertility.
Methods
Semen samples were obtained from four males with G6PD deficiency and eight healthy males as a control. In both groups, motile sperms were isolated from the seminal fluid and incubated with 0, 10, 20, 40, 60, 80, and 120 µM concentrations of H2O2. After 1 hour incubation at 37℃, sperms were evaluated for motility and viability.
Results
Incubation of sperms with 10 and 20 µM H2O2 led to very little decrease in motility and viability, but motility decreased notably in both groups in 40, 60, and 80 µM H2O2, and viability decreased in both groups in 40, 60, 80, and 120 µM H2O2. However, no statistically significant differences were found between the G6PD-deficient group and controls.
Conclusion
G6PD deficiency does not increase the susceptibility of sperm to oxidative stress induced by H2O2, and the reducing equivalents necessary for protection against H2O2 are most likely produced by other pathways. Therefore, G6PD deficiency cannot be considered as major risk factor for male infertility.
Introduction
Male factor infertility plays a role in approximately half of infertile couples. A number of etiologies have been identified as possible causes of male infertility, including genetic factors, radiation, chemotherapy, infectious diseases, ejaculatory duct occlusion, varicocele, and erectile dysfunction [1234]. Reactive oxygen species (ROS) are chemically reactive molecules and are formed as a natural consequence of the usual metabolism of oxygen, ROS plays a major role in cell signaling and homeostasis. ROS include active oxidizing agents such as H2O2, the superoxide radical, the hydroxyl radical, and singlet oxygen [5].
ROS are biologically toxic, can result in significant damage to cell structures, and are produced by two major routes. Exogenous ROS can be produced from smoke, drugs, radiation, pollutants, and similar methods. Alternatively, ROS can be generated intracellularly by several mechanisms, depending on the type of cell and tissue; however, the main sources are the seven isoforms of nicotinamide adenine dinucleotide phosphate (NADPH) oxidase that are located in the membranes, peroxisomes, mitochondria, and endoplasmic reticulum [6]. Depending on the type and concentration, ROS can have adverse or beneficial effects on sperm function. ROS are necessary for the maintenance of sperm activity and the ability of sperm cells to undergo the acrosome reaction, which allows them to penetrate the zona pellucida and fuse with the oocyte membrane [7].
Over the past decade, investigations have demonstrated that oxidative damage to sperm cells caused by ROS is one of the main reasons for impaired sperm function parameters that can result in complete loss of motility in individual sperm cells, leading to the onset of male infertility [8]. Sperm cells are very susceptible to oxidative damage due to an abundance of polyunsaturated fatty acids in the plasma membrane and a low quantity of scavenging enzymes within the cytoplasm. H2O2 can diffuse from the extracellular space across the membrane into the sperm cytoplasm [9]. The results of previous studies on antioxidant enzymes showed that H2O2 is the most toxic ROS in terms of inducing alterations in cellular function, especially in human sperm [10].
Glucose-6-phosphate dehydrogenase (G6PD) is an enzyme found in many organisms, including humans, where it is expressed in all tissues and cells and plays a key role in glucose metabolism. G6PD catalyzes the first reaction of the pentose phosphate pathway. This step converts glucose 6-phosphate to 6-phosphogluconate while also producing NADPH. One function of NADPH is the reduction of glutathione and the stabilization of catalase. Reduced glutathione and catalase play a major role in the detoxification and neutralization of ROS [11]. G6PD deficiency is the most common inborn error of metabolism, affecting an estimated 400 million people worldwide. As the G6PD gene is located on the long arm of the X chromosome (Xq28.2) and its inheritance is recessive, the disease is mostly seen in males [12].
Although the NADPH necessary for the detoxification of oxidizing agents can be produced from other pathways, we hypothesized that G6PD deficiency may decrease the NADPH and catalase content of sperm cells, thereby reducing their ability to degrade ROS. In this scenario, various types of ROS would be able to affect the sperm cells of G6PD-deficient individuals, leading to a reduction in motility and increased apoptosis due to insufficient protection against oxidative damage caused by the lower amount of reduced glutathione [13]. Thus, the objective of the present study was to study the effects of the induction of oxidative stress by H2O2 on the motility and viability of sperm cells from G6PD-deficient males in comparison to the sperm cells of healthy G6PD-normal male donors.
Methods
1. Study design
This study was conducted between November 21, 2015 and February 19, 2016 at the Motazedi Infertility Teaching Clinic of the Kermanshah University of Medical Sciences on sperm cells from the ejaculate of four males with G6PD deficiency with no known exposure to any oxidizing agents and eight healthy G6PD-normal male volunteers as the control group. Study was approved according to Iranian laws and regulations by the Institutional Review Boards of each participating teaching and referral hospital. Additionally, our study complied with the Declaration of Helsinki.
2. Fluorescent-spot test
One-milliliter blood samples were obtained from all participants in the study and collected in tubes containing ethylenediaminetetraacetic acid. A commercial kit was used to assess participants for G6PD deficiency using the fluorescent-spot test method. G6PD enzyme activity in the conversion of glucose-6-phosphate to 6-phosphogluconate and the reduction of nicotinamide adenine dinucleotide phosphate (NADH) to nicotinamide-adenine dinucleotide phosphate (reduced) NADPH2 is the basis of this qualitative method. NADPH2 exhibits fluorescence under ultraviolet light (365 nm). The presence and absence of fluorescence were considered to be indicative of active and deficient G6PD, respectively [11].
3. Basic semen analysis
Fresh semen specimens from the G6PD-deficient males and controls were collected by masturbation after 2 to 5 days of sexual abstinence and were incubated for 30 minutes at 37℃. After liquefaction, a basic semen analysis, including sperm count, motility, and morphology, was performed according to World Health Organization criteria for each sample [14]. All semen samples were evaluated by two observers to reduce interobserver variability. The discontinuous gradient method was used to isolate motile sperm cells to survey the impact of H2O2 treatment. A drop of the motile sperm cells was examined at a magnification of ×400 under a light microscope in 10 fields to record sperm motility [15].
4. Sperm viability test
Eosin-nigrosin staining was used to assess sperm viability. In summary, one drop of motile sperm cells in media was added to two drops of eosin (1.0%) in a micro tube and mixed. After 30 seconds, three drops of nigrosin (10%) were added and the solution was mixed again. After 30 seconds, a smear on a glass microscope slide was prepared from the mixture and allowed to dry in the laboratory air. Under the microscope, unstained (white) sperm cells were classified as live and differentiated from pink-stained sperm cells, which were classified as dead. For each smear, at least 200 sperm cells were evaluated, and finally, the percentage of unstained sperm cells was considered to be the percentage of viable sperm [16].
5. H2O2 treatment
After isolation of motile sperm from semen samples using the discontinuous gradient method, 50 µL from the motile sperm suspension of each group was added to 950 µL of Ham's F10 medium containing H2O2 (Sigma-Aldrich, Taufkirchen, Germany) in order to create solutions with final concentrations of 0, 10, 20, 40, 60, 80, and 120 µM H2O2, and the solutions were incubated for 1 hour at 37℃. At the end of the incubation time, samples were centrifuged, the supernatant was removed, the sperm concentration was adjusted to 30×106/mL, and the samples underwent a sperm motility and viability analysis [1417].
6. Statistical analysis
SPSS ver. 16.0 (SPSS Inc., Chicago, IL, USA) was used for the statistical analysis. The chi-square test was used to characterize the relationships between dependent and independent variables. The p-values <0.05 were considered to indicate statistical significance. The mean and standard deviation were calculated for each variable.
Results
In the basic semen analysis, all parameters, including volume, count, motility, morphology, and viability, were within the normal range in all participants in both groups (G6PD-deficient males and healthy controls) (Table 1).
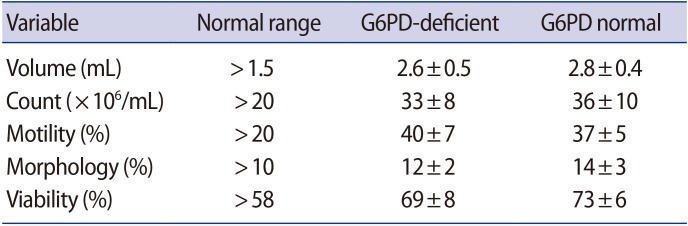
Results of the basic semen analysis of G6PD-deficient and G6PD-normal males
1. Sperm motility
The results of the sperm motility test in G6PD-deficient males and the control group (after discontinuous gradient purification) in cells subjected to different concentrations of H2O2 are shown in Figure 1. Very moderate decreases in sperm motility were observed in the 10 and 20 µM concentrations of H2O2, while motility was found to dramatically drop at 40, 60, and 80 µM concentrations of H2O2 in both groups. Motility was completely eliminated in the 120 µM concentration of H2O2. However, no statistically significant differences in motility were found between the G6PD-deficient group and the normal control group after exposure to different concentrations of H2O2 (p>0.05).

Effects of different concentrations of H2O2 on sperm motility. G6PD, glucose-6-phosphate dehydrogenase.
2. Sperm viability
The impact of different concentrations of H2O2 on sperm viability (after discontinuous gradient purification) is shown in Figure 2. A very small decrease in sperm motility was observed at 10 and 20 µM concentrations of H2O2 in both groups (G6PD-deficient males and healthy controls) (p>0.05). A sharp decrease in the viability of sperm cells after treatment with H2O2 concentrations higher than 40 µM was seen in both groups. Overall, no statistically significant differences were found in motility between the G6PD-deficient group and healthy controls after treatment with different concentrations of H2O2 (p>0.05).
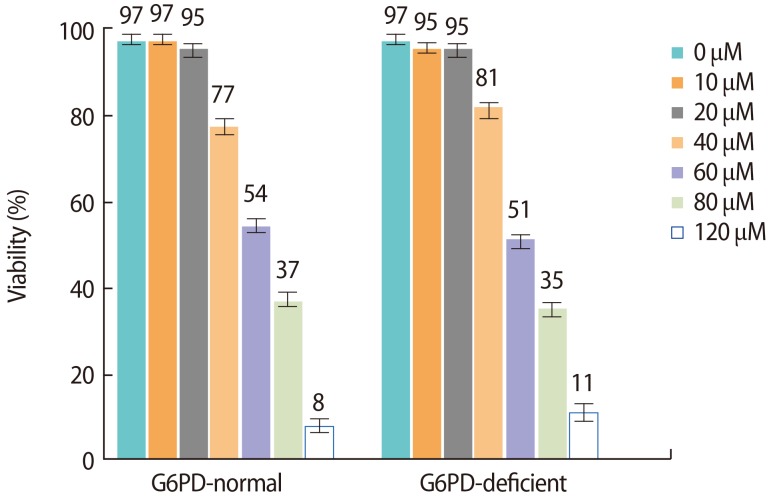
Effects of different concentrations of H2O2 treatment on sperm viability. G6PD, glucose-6-phosphate dehydrogenase.
Discussion
In the present study, we found gradual decreases in the motility and viability of sperm cells after exposure to different concentrations of H2O2 in an artificial medium. H2O2 concentrations up to 20 µM showed little effect on motility and viability; however, motility was the first parameter to be affected by higher concentrations of H2O2. Few studies have yet been performed on the effects of H2O2 on the structure and function of sperm cells under different concentrations and laboratory conditions [141718]. Schraufstatter et al. [18] showed that H2O2 concentrations of up to 50 µM did not damage cellular functions. They also reported that the loss of forward progressive motility within a very short time was observed in sperm cells treated with 600 µM or greater concentrations of H2O2.
ROS have been hypothesized to play a causative role in the etiology of sperm dysfunction. Several mechanisms have been proposed to explain this effect. Peroxidation of the unsaturated fatty acids within the sperm plasma membrane results in loss of the fluidity, structure, and function of the membrane. ROS also cause a decrease in the rate of glycolysis by inactivating adenylate dehydrogenase and reducing the amount of lactate available [78]. Another important factor is the effect of ROS on the mitochondria, which produce adenosine triphosphate (ATP). ATP is a usable form of energy for the cell and, in the case of sperm cells, sustains their motility, which represents one of the main determinants of male fertility [19].
Oxidative phosphorylation is a process in which ATP is produced by ATP synthase using energy provided by proton transfer in the electron transport chain across the inner mitochondrial membrane. In normal conditions, oxygen is reduced and water is produced; however, in approximately 0.2% to 2% of electrons passing through the electron transport chain, oxygen is reduced incompletely and ROS are produced, which can cause damage to the cell. If significant damage accumulates in the mitochondria, apoptosis occurs [20]. Mitochondrial metabolism is crucial for several functions, especially regarding the motility of sperm cells. Additionally, the mitochondria are a site of the major metabolic pathways during the development of germ cells and fertilization [21].
In our study, although H2O2 as an oxidative stress agent affected the motility and viability of sperm and decreased these parameters at high concentrations, no significant differences were found in G6PD-deficient patients compared to the control group (Figures 1, 2). Some studies on associations between G6PD deficiency and susceptibility to oxidative stress have been performed [2223]. Efferth et al. [22] performed a study in Germany on the impact of H2O2 on mononuclear cells in G6PD-deficient patients. They concluded that the defective detoxification of ROS by glutathione may cause increased DNA damage, leading to an increased apoptosis rate in G6PD-deficient mononuclear cells. Tian et al. [23] examined the role of G6PD in the regulation of cell death using cell lines. The cell lines were incubated with 50 µM and 100 µM H2O2 in the presence and absence of known G6PD inhibitors. They showed that changes in G6PD activity affected oxidative stress-induced cell death and apoptosis, and concluded that G6PD plays an important role in cell death by regulating intracellular oxidation-reduction conditions.
Although some studies related to sperm dysfunction in the presence of oxidative stressors, such as H2O2, have been previously published, this appears to be the first report in which the role of G6PD enzyme deficiency in increased susceptibility to oxidative stress was investigated in the sperm cells of males affected by this disorder [141724].
Cell damage arises when the balance between the production and scavenging of ROS is disturbed, and such imbalances have been shown to be associated with idiopathic male infertility [8]. The detoxification of H2O2 depends on NADPH availability, which provides the reducing equivalents required for oxidation-reduction reactions involved in protecting against the toxicity of ROS. In the majority of body cells in humans, the main source of NADPH is the pentose phosphate pathway [6].
However, our results suggest that the pentose phosphate pathway does not play a major role in the production of NADPH and the detoxification of ROS in sperm cells, although more studies with larger samples should be performed in order to confirm these results. Sperm cells have many mitochondria in locations that play a vital role in their functions, especially in motility of the flagellum [25]. However, several other alternative mechanisms exist for the generation of NADPH, and almost all of them require the presence of mitochondria [13]. The mitochondria of sperm are different from the mitochondria of other somatic cells in terms of metabolism and morphology. These metabolic differences are related to the presence of certain enzyme isoforms, which are characterized by particular regulatory mechanisms and kinetic characteristics [25]. The key enzymes in these mechanisms are NADPH-linked malic enzyme, NADPH-linked isocitrate dehydrogenase, isocitrate dehydrogenase and glutamate dehydrogenase, nicotinamide nucleotide transhydrogenase, and NADH kinase, which produces NADPH and adenosine diphosphate, using NADH and ATP as substrates [26].
It seems that one or more of these alternative pathways may play a major role in the detoxification of ROS in sperm cells. Finally, we speculate that the sperm cells of G6PD-deficient males are likely to be much more susceptible to DNA damage caused by oxidative stress than the sperm cells of G6PD-normal individuals. The impact of H2O2 on sperm DNA was not investigated in this study, but results of previous studies have shown that H2O2 causes DNA damage in cells [2227]. Thus, further studies are recommended on the relationship between G6PD deficiency and sperm DNA damage.
In conclusion, based on our study, sperm cells of G6PD-deficient males are not more susceptible than those of G6PD-normal males to oxidative stress induced by H2O2. It seems that the pentose phosphate pathway is not the main source of the reducing equivalents necessary for protection against the toxicity of ROS in sperm and that these reducing equivalents are most likely produced by other pathways. Studies should be performed on the relationship between G6PD deficiency and sperm DNA damage in order to rule out any potential role of G6PD enzyme deficiency in sperm dysfunction and subsequent male infertility.
Acknowledgments
The authors would like to thank the members of Motazedi Infertility Clinic Laboratory for their constant support.
Notes
Conflict of interest: No potential conflict of interest relevant to this article was reported.