Genomic aspects in reproductive medicine
Article information

Abstract
Infertility is a complex disease characterized by extreme genetic heterogeneity, compounded by various environmental factors. While there are exceptions, individual genetic and genomic variations related to infertility are typically rare, often family-specific, and may serve as susceptibility factors rather than direct causes of the disease. Consequently, identifying the cause of infertility and developing prevention and treatment strategies based on these factors remain challenging tasks, even in the modern genomic era. In this review, we first examine the genetic and genomic variations associated with infertility, and subsequently summarize the concepts and methods of preimplantation genetic testing in light of advances in genome analysis technology.
Genetic causes of infertility
It is estimated that approximately 50% of infertility cases result from genetic defects at various levels, from chromosome abnormalities to single nucleotide variations [1]. However, aside from numerical sex chromosome abnormalities, the genetic causes of infertility are highly heterogeneous, making it challenging to understand the genetic background of individual patients. While advanced genome analysis methods, such as chromosome microarray (CMA) and next-generation sequencing (NGS), are commonly used for many genetic diseases, only conventional chromosome analysis and Y chromosome microdeletion tests are currently offered as routine genetic tests for infertile couples. In this section, we briefly discuss the well-known genetic causes associated with infertility.
1. Chromosome abnormalities in infertile couples
Numerical and structural chromosome abnormalities are well-established causes of infertility, accounting for up to 20% of cases in infertile males and 7.82% in infertile females [2,3]. Therefore, chromosome analysis using peripheral blood is typically recommended for new patients. Klinefelter syndrome (47, XXY) and Turner syndrome (45, X) are the most common causes of infertility in males and females, respectively [4,5]. Klinefelter syndrome affects 0.1% to 0.2% of men, while Turner syndrome is found in approximately 0.04% of women [6,7].
Balanced structural chromosome abnormalities, such as translocations and inversions, are a well-established cause of infertility. Although carriers may exhibit little to no clinical phenotype, an unbalanced chromosome can result from unusual pairing and recombination between homologous chromosomes. This unbalanced chromosome may be passed onto the fetus, leading to a higher rate of miscarriages [8,9]. A Robertsonian translocation (RT), a condition where the long arms of two acrocentric chromosomes attach to each other, occurs in approximately 1 to 1.2 out of every 1,000 newborns. The most common form of RT is rob(13;14), accounting for 75% of all RT cases. Carriers of this form are known to have a high likelihood of male infertility and recurrent pregnancy loss [10].
2. Chromosomal abnormalities in gametes and early embryos
Nondisjunction can occur in either homologous chromosomes (during meiosis I) or sister chromatids (during meiosis II), leading to gametes that either lack a chromosome or possess an extra copy of one. Most autosomal aneuploidies found in fetuses are primarily of maternal origin, resulting from nondisjunction in meiosis I. It is well-established that chromosomal nondisjunction during oogenesis is age-related, with a particularly sharp increase observed in women aged 35 and older. This can be attributed to the unique developmental process in oocytes, which begins before birth and concludes after fertilization [11,12]. As oocytes age, their capacity for microtubule formation and dissociation during anaphase diminishes. This can lead to chromosomal nondisjunction, thereby increasing the risk of oocyte aneuploidies, implantation failure, or spontaneous abortion [13,14].
Advanced paternal age can also impact fertility. While there have been reports of increased aneuploidy in sperm, the effect is not as pronounced as it is in females. Instead, age may lead to decreased testicular function, sperm DNA damage, and an increase in mutations [15,16]. It is noteworthy that in patients with oligozoospermia and teratozoospermia, who have reduced sperm quality, a higher frequency of sperm aneuploidies was observed, regardless of age [17-19]. Furthermore, chromosome abnormalities in cleavage-stage in vitro fertilization (IVF) embryos are known to be common. Numerous studies have found that more than 50% of embryos at this stage exhibit chromosome abnormalities [20,21]. Vanneste et al. [22] demonstrated that mitotic nondisjunction and chromosome instability are common in human cleavage-stage embryos. They found that even within a single embryo, each blastomere can have different types of numerical and structural chromosome abnormalities [22]. This is a major factor underlying the reduced effectiveness of preimplantation genetic screening (PGS) and the difficulty in determining which embryos are suitable for transplantation.
Intracytoplasmic sperm injection (ICSI) was introduced in the 1990s as a treatment for infertile patients who had experienced unsuccessful IVF attempts [23,24]. Although ICSI has been proven effective for treating male infertility, concerns about the safety of offspring have been raised due to the invasiveness of the procedure [25]. Early studies published shortly after the introduction of ICSI reported an increased risk of chromosomal abnormalities in fetuses [8,26,27]. However, a recent systematic review found no statistically significant association between ICSI or IVF and fetal aneuploidy [25,28]. Despite these concerns, ICSI and IVF continue to be routinely performed in many clinics and remain effective treatment methods for infertile couples.
3. Epigenetic abnormalities
Epigenetic modifications refer to inheritable changes that can manifest differently due to environmental factors, without any alteration to the DNA sequence [29]. These modifications are recognized as a crucial factor in regulating gene expression across a range of biological processes, including gametogenesis [30]. For example, alterations in modifications such as histone methylation, changes in the P1/P2 ratio, and methylation patterns in imprinting genes may lead to infertility by impacting the maintenance of pregnancy and fetal development [31].
As primordial germ cells migrate to the gonadal ridge, most DNA methylation patterns, which were previously programmed similarly to somatic tissue, are soon erased and subsequently re-established. This reprogramming process allows the correction of heritable methylation errors and introduces new methylation patterns. However, changes in the expression of essential genes due to either hyper- or hypo-methylation can disrupt gametogenesis [30]. For instance, studies have reported that infertile patients with low sperm concentration, motility, and morphology exhibited higher levels of methylation in their sperm than healthy individuals [32].
In addition to methylation, acetylation and ubiquitination have been demonstrated to play a role in modifying histone proteins and regulating gene expression, which is implicated in infertility. The transition from histone to protamine is necessary for the proper packaging of the required DNA into haploid spermatids. This transition is facilitated by the hyperacetylation of histone H4, which reduces the affinity of histone H4 for DNA [30]. Sonnack et al. [33] observed a decrease in the hyperacetylation of histone core proteins in infertile patients, suggesting the importance of these modifications in the transition from round spermatids to mature, fertile spermatozoa.
4. Copy number variations
Copy number variations (CNVs) are submicroscopic chromosomal structural changes involving alterations of 1 kilobase or more [34]. CNVs are known to originate from chromosomal structural rearrangements, including inversion, translocation, and deletion/insertion. The clinical significance of a CNV is determined by the size of the variation and the specific genes within the affected region [31].
A CNV variant associated with infertility is the microdeletion of the azoospermia factor (AZF) region on the Y chromosome, which accounts for approximately 10% of all infertile male patients [35,36]. Deletions within each segment of the AZF region are known to present a range of infertility symptoms, from severe oligozoospermia to Sertoli-cell-only syndrome [37]. Conversely, CNVs identified on the X chromosome are considered a primary cause of female infertility, including conditions such as premature ovarian failure [38,39]. However, many of the CNVs discovered in these studies are still classified as variants of uncertain significance, indicating a need for further research to understand their actual impact.
The CMA technique enables rapid, automated detection of structural abnormalities or submicroscopic CNVs on targeted chromosomes at units of 4 to 10 Mb or less [40,41]. First employed in reproductive studies in 2005, CMA was used to analyze the causes of miscarriages attributed to chromosomal abnormalities, as well as fetuses exhibiting morphological abnormalities [40,42,43]. Due to its effectiveness in cases that could not be karyotyped, CMA is now recommended for ongoing pregnancies, as well as cases involving congenital anomalies and fetuses with morphological abnormalities [40].
Recently, CMA has been introduced as an advanced method for detecting more precise sizes and breakpoints of Yq microdeletions. Zhu et al. [44] identified various CNVs of the azoospermia factor c (AZFc) regions in patients with azoospermia and severe oligozoospermia. This suggests that the genes located in these regions (DAZ1, DAZ2, DAZ3, CDY1, and BPY2) may play a significant role in male infertility [44]. CMA can provide more detailed information on chromosomal CNVs than traditional karyotyping can. However, it does have limitations, such as its inability to detect balanced translocations or low-level mosaicism. As a result, there has been a shift towards more advanced technology, NGS, allowing more sophisticated and accurate detection of chromosomal variations.
Single gene defects
Hundreds of genes associated with infertility have been identified through mouse models. These genes play a role in sex determination, the formation of reproductive organs, gametogenesis, cell division, and the regulation of pituitary and gonadal functions within the endocrine system [45]. While pathogenic variants in human homologs of some of these genes have been discovered in patients with infertility, the application of sequencing analysis as a clinical test has not been widely adopted.
For female infertility, genetic analysis of fragile X mental retardation 1 (FMR1) and anosmin 1 (ANOS1) (also called KAL1) has been recommended for patients with premature ovarian insufficiency (POI) and hypogonadotropic hypogonadism (HH), respectively, since the early 2000s [46]. Early research identified variants in genes encoding gonadotropic hormones such as gonadotropin-releasing hormone, follicle-stimulating hormone, and luteinizing hormone, as well as their receptors [11,47]. In addition, other POI-related genes, such as FOXL2, SOHLH1, GDF9, NOBOX, STAG3, and TUBB8, have been found to affect oocyte maturation and meiosis [48-53]. Polycystic ovary syndrome (PCOS), another major cause of reproductive problems, is the most common endocrine-metabolic disorder in reproductive-aged women [54]. Patients with PCOS often show subfertility due to ovulatory dysfunction [55]. Numerous genes associated with PCOS have been identified through genome-wide association studies (GWAS), and these genes play roles in gonadotrophin action, ovarian follicle development, diabetes, and organ growth [56]. few clear pathogenic variants in these genes have been identified, and genetic testing for diagnosis is not typically conducted.
Since the mid-2010s, studies have been undertaken to screen fertility-related genes simultaneously using NGS technologies [57,58]. Volozonoka et al. [59] proposed a diagnostic gene panel for female reproductive failure, based on a systematic review of previously published papers. This gene panel includes 79 genes associated with conditions such as HH, ovarian dysgenesis, premature ovarian failure/insufficiency, oocyte maturation, and fertilization failure [59]. Based on these studies, it is anticipated that targeted-exome analysis could be introduced in the near future as a diagnostic tool for women with fertility disorders.
Similar to female infertility, the etiology of male infertility is also heterogeneous. Alongside advancements in NGS technology, studies have been conducted to identify new potential genes associated with male infertility. Over 3,500 papers have been published on this topic, suggesting the involvement of hundreds of genes. However, diagnosis is currently limited to karyotype, AZF microdeletion, and cystic fibrosis transmembrane conductance regulator (CFTR) analysis [60].
The primary phenotypes of male infertility encompass both oligozoospermia and azoospermia, which are defined by a diminished sperm count or a complete lack of sperm production. Additionally, asthenozoospermia and teratozoospermia are included, both of which are distinguished by irregularities in sperm motility and morphology, respectively [61]. Well-known candidate genes for male infertility, especially for non-obstructive azoospermia (NOA), include TEX11, SYCP3, and SYCE1, which are known to be involved in meiosis [35,62]. Furthermore, the genes that encode proteins of the axonal dynein cluster, such as DNAH1, DNAH5, and DNAH11, have been suggested to be associated with asthenozoospermia. Variants of AURKC and SPATA16 were also identified in patients showing sperms with multiple flagella or globozoospermia [61,63]. Since all genes involved in spermatogenesis are candidate genes for male infertility, basic research on the gene-disease relationship for each gene is very important.
A systematic review published in 2018 integrated data from previous studies and recent NGS analyses to suggest 92 candidate genes that have at least moderate evidence supporting their involvement in male infertility [60]. This indicates potential diagnostic advancements in the field of male reproduction. However, this list only includes approximately 18% of gene-disease relationships, suggesting that a significant number of associations still require substantial evidence. This limitation may stem from the nature of medical genetics, which primarily focuses on inherited variations. For instance, many genes that result in non-syndromic phenotypes exhibit recessive inheritance, which is only applicable in specific cases, such as those involving consanguineous families. Moreover, the exploration of associations with autosomal dominantly inherited causative genes of male infertility is limited unless a de novo variation occurs, as the transmission of variation to subsequent generations is rare. This is evidenced by the fact that only three genes (DMRT1, KLHL10, and SYCP3) that exhibit autosomal dominance have moderate evidence among the 92 genes on the list. To address these limitations, future research on male infertility should include repeated reanalysis of genetic data, as well as more in-depth functional and cohort studies.
Preimplantation genetic testing
With advances in genome analysis technology, preimplantation genetic testing (PGT) methods have also changed (Figure 1). The early methods, which were labor-intensive, required high skill, and were time-limited, are now being replaced by more automated and high-throughput methods. PGT can be categorized into several types based on the nature of the genetic defects (PGT for aneuploidy [PGT-A], PGT for structural rearrangement [PGT-SR], PGT for monogenic disorder [PGT-M], PGT for polygenic disorder [PGT-P]) and the method of obtaining genetic materials (PGT from embryo biopsy and non-invasive PGT [niPGT]). In this paper, our primary focus is on the technical aspects of PGT.
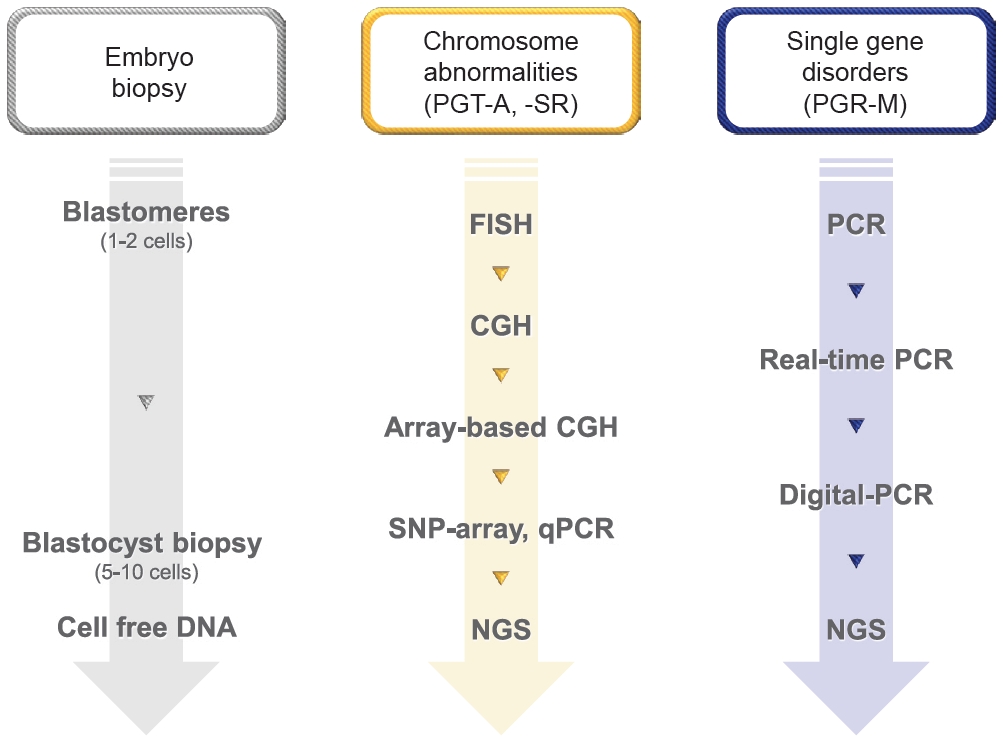
Technical advances for preimplantation genetic testing (PGT). PGT-A, PGT for aneuploidy; PGT-SR, PGT for structural rearrangement; PGT-M, PGT for monogenic disorder; FISH, fluorescent in situ hybridization; CGH, comparative genomic hybridization; SNP, single nucleotide polymorphism; qPCR, quantitative polymerase chain reaction; NGS, next-generation sequencing; PCR, polymerase chain reaction.
1. Preimplantation genetic testing for aneuploidy
PGT-A, previously known as PGS, is a genetic test used to determine if embryos created through IVF have genomic imbalances, including chromosomal aneuploidies. Munne et al. [64] first reported PGT-A in 1993. From then until the early 2000s, one to two blastomeres were typically isolated from 8-cell stage embryos. Subsequently, five to nine chromosomes, most commonly associated with fetal chromosomal aneuploidies, were primarily analyzed using fluorescent in situ hybridization (FISH) [64]. However, the technical limitations of FISH, such as its inability to identify aneuploidy in all chromosomes, coupled with the high mosaicism rate of 8-cell embryos, significantly reduced the accuracy of PGS [65].
As advancements have been made in embryo culture methods and genome analysis technology, the use of PGT-A, which incorporates trophectoderm (TE) biopsy, vitrification, and array-based genome analysis or NGS techniques, has become commonplace [66,67]. TE biopsy offers several advantages over blastomere biopsy. By isolating more cells (typically 5 to 10), the failure rate of DNA amplification can be reduced, leading to more reliable diagnostic results. Furthermore, TE biopsy is less invasive to embryos than blastomere biopsy [68]. Array-based comparative genomic hybridization (array CGH) enables the identification of segmental imbalances in chromosomes, as well as aneuploidies of all 24 human chromosomes. Initially, array CGH was used to detect genomic imbalances in patients exhibiting abnormal phenotypes or in tumors. It was later introduced for genetic screening of preimplantation embryos [69-72].
NGS technologies are presently the most frequently utilized for PGT-A. This is due to the commercial availability and cost-competitiveness of NGS-chips for PGS and standard analysis protocols (VeriSeq, Illumina; ReproSeq, Thermo Fisher) provided by centralized service providers. These technologies offer a higher resolution compared to earlier array-based techniques, enabling more accurate identification of mosaicism with a 20% to 80% ratio of normal-to-abnormal cells [73]. However, this advantage often leads to challenges in determining which embryos are suitable for transplantation.
Several researchers have argued that PGS may not be effective in improving the actual fertility rate [74,75]. These authors concur that the rates of aneuploidy and miscarriage can be diminished by screening embryos using PGS, but only for patients who have a sufficient number of embryos for analysis and a favorable prognosis. Conversely, they also highlighted the potential for discarding embryos that could potentially develop into healthy newborns in less favorable cases. For example, Scriven [76] noted that while PGT-A is a superior test for older individuals in terms of reducing miscarriage risk, their cumulative ongoing pregnancy rate is lower than that of younger individuals. Furthermore, Greco et al. [77] found that approximately 30% of implanted mosaic embryos developed into euploid newborns. A recent cellular study demonstrated that a small proportion of euploid cells could salvage mosaic embryonic organoids (gastruloids) when combined with aneuploid cells, suggesting a potential in vivo mechanism for aneuploid elimination in humans [78]. Nevertheless, the efficacy of PGS remains a topic of ongoing debate.
2. Preimplantation genetic testing for monogenic disorders
PGT-M, formerly termed preimplantation genetic diagnosis (PGD), is a test used to determine if preimplantation embryos are at risk for certain Mendelian monogenic diseases. PGD, when administered to carriers of balanced structural chromosome abnormalities, is now classified separately as PGT-SR. The procedures and testing methods for PGT-SR are essentially the same as those for PGT-A and PGT-M. PGT-M can be administered to couples who are at risk of transmitting a specific genetic disorder to their offspring, or to couples who are both carriers of an autosomal recessive disease.
In 1990, the first instance of PGD was reported for an X-linked genetic disorder, achieved through sexing via the amplification of Y chromosome-specific sequences [79]. Initially, PGT-M was mainly performed using polymerase chain reaction (PCR)-based methods [80]. Given that only a minuscule amount of DNA is available, PCR-based techniques were prone to errors due to amplification failure, allele dropout (where only one of two alleles is randomly amplified), and contamination, among other issues [81]. These limitations of PCR-based methods have been significantly mitigated through the advent of whole genome amplification technologies and the analysis of multiple loci linked to the target sequence using short tandem repeat (STR) markers [82-84]. Since the mid-2010s, reports of PGT-M using NGS-based technology have emerged [85,86]. This method can simultaneously analyze chromosome copy number and pathogenic variants.
Karyomapping, a technique frequently employed in PGT-M, involves genome-wide linkage analysis for a wide variety of pathogenic variants, ranging from CNVs to single gene defects, including imprinting disorders [87]. This method analyzes hundreds of thousands of single nucleotide polymorphisms (SNPs), which are evenly distributed across the entire genome, thereby reducing the potential for diagnostic errors. Furthermore, it eliminates the need to select STR markers for specific genetic diseases, making it possible to standardize the method for nearly any disease.
3. Preimplantation genetic testing for polygenic disorders
Treff et al. [72] proposed PGT for polygenic conditions such as type 1 and type 2 diabetes, coronary artery disease, and certain types of cancers. However, it has not been incorporated into medical care due to challenges in interpreting results and the ethical issues it raises. The results of PGT-P are determined as the relative risk of disease based on the polygenic risk score for numerous involved genes, rather than a straightforward distinction between normal and affected. These polygenic risk scores are obtained through GWAS of a population, and thus may vary among different populations [88]. Typically, the variants associated with diseases identified through GWAS exhibit a low odds ratio, less than 1.2. Consequently, the predictability of PGT-P in selecting embryos for transfer is low. Furthermore, a chosen embryo may display conflicting disease risks for several different polygenic diseases, being favorable for one disease and at risk for another. Therefore, a balance of disease risks must be considered when selecting embryos [89]. Another important ethical question is whether it is acceptable to select embryos for treatable and preventable diseases. Currently, it seems that there are numerous technical and ethical issues that need to be addressed before PGT-P can be implemented.
4. Non-invasive preimplantation genetic testing
Traditional methods of biopsy for DNA extraction pose a risk of causing harm to embryos. To mitigate this risk, niPGT methods that utilize blastocoel fluid and spent culture medium (SCM) have been developed. The aspiration of blastocoel fluid is a routine procedure performed before the vitrification of embryos. Palini et al. [90] were the first to demonstrate the feasibility of PGT using cell-free DNA in blastocoel fluid (BF-DNA). They found approximately 10 pg of BF-DNA and successfully amplified several gene-specific sequences. They also performed array CGH using whole genome amplification of BF-DNA. However, array CGH results were only obtained in two out of five cases. Since then, numerous studies on niPGT using BF-DNA have been reported. More recent studies have shown an increase in the efficiency of whole genome amplification and the validity of array CGH results [91,92]. In addition, Zhang et al. [93] showed that whole genome amplification products obtained from BF-DNA could be used for PGT-M.
SCM serves as an additional source of circulating free DNA for niPGT-A. Hammond et al. [94] found that SCM contained higher levels of genetic material originating from the embryo, and that the quantity of DNA increased during the course of embryo culture. However, they concluded that PGT using DNA from SCM was not suitable due to the limited amount of DNA and the contamination from maternal DNA [94]. Subsequent reports have indicated a more promising potential for niPGT-A using SCM. In instances of 24-hour culture of thawed blastocysts, niPGT-A proved to be more reliable than PGT-A following a TE biopsy [92,95,96]. The most recent studies have demonstrated the increased clinical applicability of niPGT-A using SCM. This has been achieved through the modification of culture methods, the enhancement of genome amplification tools, and the selection of embryos based on morphological grading [97-99].
Mosaicism remains a factor that compromises reliability, even in the context of niPGT-A. Many cases of mosaic chromosome aneuploid or segmental aneuploid are often revealed to be false positive. Cai et al. [100] employed morphological assessment to select and transfer high-quality embryos. Their research demonstrated that over 90% of mosaic aneuploids identified in niPGT-A were actually euploid, constituting false positives [100]. The niPGT-M method, which utilizes BF-DNA and SCM, appears to present more challenges than niPGT-A. While there have been instances of using BF-DNA and SCM to amplify certain targeted genes or analyze disease-associated SNPs, significant variations in amplification rates and concordance with TE biopsy results have been observed [101,102].
Considering the results of studies concerning niPGT to date, their clinical significance is challenging to evaluate due to the small sample sizes and variations in methodologies. There have been several clinical trials related to niPGT, and a relatively large, multicenter, double-blind, randomized controlled trial is currently underway to assess the clinical relevance of niPGT [103].
Expanded carrier screening in reproductive medicine
As NGS has become more common, it has enabled carrier testing for a multitude of genetic diseases simultaneously. Approximately 2% to 4% of couples planning to conceive are at risk of having a child with an autosomal recessive disorder or an X-linked disorder [104]. Traditionally, carrier screening has been conducted on adults who have a family history of a specific genetic condition. However, such genetic conditions are typically rare, and most affected children are born to couples with no known family history [105]. Several scientific societies, including the American College of Obstetrics and Gynecology (ACOG), the American College of Medical Genetics and Genomics (ACMG), and the European Society of Human Reproduction and Endocrinology (ESHRE), have proposed the concept of expanded carrier screening (ECS). ECS is a preconception genetic screening for common and severe recessive diseases, intended for healthy individuals with no known family history of these conditions. These societies have also suggested a list of genes to be included in the gene panel [106-109]. Although ECS has many advantages, such as the simultaneous analysis of multiple genetic diseases, cost-effectiveness, and the provision of reproductive planning for at-risk couples—including the avoidance of genetic diseases through PGT—a careful approach is required when considering its routine implementation, particularly in Korea. Beyond technical limitations, such as difficulties in interpreting results, the potential for incidental findings, and variants of unknown significance, there are also considerations regarding whether the disease or mutation is suitable for PGT. Furthermore, a societal consensus, including agreement within the medical community, is necessary for the implementation of ECS [110].
Conclusion
As genome analysis technologies advance, they enable the simultaneous analysis of a spectrum from single nucleotide variants to large CNVs. Consequently, all types of PGT, including PGT-A, -SR, and -M, can be performed concurrently on a single platform. Furthermore, the application of more sophisticated genome analysis techniques, such as single-cell sequencing, to PGT could simplify and enhance the accuracy of determining whether embryos carry disease-causing variants. As we continue to uncover more genetic causes and risk factors for infertility, it becomes possible to prevent infertility or boost the success rate of IVF through patient-specific treatment. Gene editing technology, utilizing the Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR)-Cas9 system, has the potential to correct mutations in germline cells, gametes, and even zygotes [111,112].
The genome era has greatly influenced the field of infertility, bringing about numerous positive effects. However, the application of currently developed technologies to clinical practice presents a host of technical and ethical issues. These challenges necessitate a consensus within both the medical community and society at large.
Notes
Conflict of interest
No potential conflict of interest relevant to this article was reported.
Author contributions
Conceptualization: MG, SHS.
Data curation: MG, SHS.
Formal analysis: MG, SHS.
Funding acquisition: SHS.
Methodology: MG, SHS.
Project administration: SHS.
Visualization: MG, SHS.
Writing-original draft: MG, SHS.
Writing-review & editing: MG, SHS.