Introduction
Cholesterol is a sterol biosynthesized in all animal cells due to its structural role in animal cell membranes [1]. In addition, cholesterol is a precursor to the biosynthesis of steroid hormones, bile acids, and vitamin D [2]. Most cholesterol synthesis takes place in the liver, although some occurs in the kidneys, intestines, and adrenal glands [3]. Hypercholesterolemia is associated with male reproductive dysfunction. For example, a high-cholesterol diet in male rats is associated with decreases in sperm quality and in the nuclear dimensions of Leydig cells, both of which lead to infertility [4,5].
Probiotics are live microbial food additives that have beneficial effects on host health by regulating the microbial balance of the intestinal tract. The most widely used probiotic microorganisms are Bifidobacterium and Lactobacillus species. Lactic acid bacteria cultures, especially dairy products containing Lactobacillus acidophilus, have been found to lower serum cholesterol and increase the quantity of fecal Lactobacilli [6].
Differing opinions exist regarding the mechanism of the serum cholesterol-lowering effects of probiotic bacteria. The most common belief is that probiotics are rapidly eliminated from the intestinal tract by breaking down bile salts into free acids. The synthesis of bile acids from cholesterol decreases the overall concentration of cholesterol in the body, as free bile salts are excreted from the body [7]. Globally, hypercholesterolemia is a very common health issue. In this study, we investigated the addition of the probiotic L. acidophilus to male rats fed a high-cholesterol diet. We aimed to investigate the histopathological impact on testicular tissue along with the effects on testosterone (TES), follicle-stimulating hormone (FSH), luteinizing hormone (LH), androgen-binding protein (ABP), and apoptosis.
Methods
1. Animal materials
Twenty-four male Sprague-Dawley rats 10ŌĆō12 weeks of age and weighing 300ŌĆō350 g were used in this study. The rats were obtained from Ondokuz Mayis University Experimental Animals Application and Research Center. The present study was conducted with the permission of the Ondokuz May─▒s University Animal Experiments Local Ethics Committee (2016/27). During the study, an environment with a room temperature of 22┬░C┬▒2┬░C, 60% humidity, and a 12/12 hour light/dark cycle was provided. The experimental animals were fed ad libitum throughout the study.
2. Preparation of probiotic suspensions
Lyophilized L. acidophilus ATCC 4356 was supplied for use as a probiotic medication. The bacteria were inoculated with de Man, Rogosa, and Sharpe (MRS) agar for viability, and the purity was confirmed by diluting the lyophilized bacteria with MRS broth. One milliliter of the culture, determined to be viable and pure, was added to 500 mL of MRS agar and incubated at 35┬░C for 48 hours. At the end of the incubation period, the suspension was inoculated to MRS agar plates and incubated at 35┬░C for 48 hours. After incubation, the bacterial colonies were counted, and the concentration of bacteria (colony-forming units [CFU]/mL) in the main culture was calculated. Then, 1010 CFU/mL bacteria were suspended in the main culture, and a probiotic suspension of L. acidophilus was prepared for use as treatment [8].
3. Animal experiment
For the animal experiment, three groups of eight rats were established. (1) Group 1 (n=8, control group [C]): the animals of the control group were fed standard rat food ad libitum for 8 weeks. (2) Group 2 (n=8, 2% cholesterol [HC]): this group was fed standard rat food with 2% cholesterol ad libitum for 8 weeks [9,10]. (3) Group 3 (2% cholesterol+L. acidophilus [HCL]): this group was fed standard rat food with 2% cholesterol ad libitum for 8 weeks [9,10]. For the last 4 weeks of the experiment, L. acidophilus probiotic (2├Ś108 CFU/mL/day) was administered via oral gavage [8].
At the end of the study, the rats were individually weighed, and 10% ketamine (Ketasol; Richter Pharma Ag, Wels, Austria; 0.8ŌĆō1.3 mL/kg) and 2% xylazine (Basilazine; Bavet, Istanbul, Turkey; 2ŌĆō5 mg/kg) were applied via intraperitoneal injection. Each rat was decapitated after the blood was drained from the heart, and testicular tissue samples were taken after necropsy.
4. Preparation of testicular tissue samples for enzyme-linked immunosorbent assay
The testicular tissue samples were weighed after washing with ice-cold phosphate-buffered saline (0.01 mol/L, pH 7.0ŌĆō7.2) before homogenization. After the tissue was divided into small pieces, homogenization was performed in 5ŌĆō10 mL phosphate buffer solution with an ice homogenizer. The cell membrane was subsequently broken down twice with ice ultrasonics, and ice cream thawing was applied twice for better cell disintegration. The homogenates were centrifuged at 5,000 ├Śg (4┬░C) for 5 minutes. They were then divided into supernatant aliquots and stored at ŌłÆ80┬░C until the analyses were performed.
5. Determination of testicular TES, FSH, and factor-associated apoptosis levels using enzyme-linked immunosorbent assay
Specific enzyme-linked immunosorbent assay (ELISA) kits were used to assess the levels of TES, FSH, and factor-associated apoptosis (FAS) in the testicular tissue supernatants of each rat. The ELISA procedures were performed according to the manufacturerŌĆÖs instructions, and the absorbance levels (as optical density values) of the ELISA plates were measured with an ELISA reader.
6. Determination of serum biochemical parameters
The LH and ABP levels of each rat were determined using specific ELISA kits. The ELISA procedures were performed according to the manufacturerŌĆÖs instructions, and the absorbance levels (as optical density values) of the ELISA plates were measured with an ELISA reader. Serum TAC levels were measured spectrophotometrically with an autoanalyzer. For this analysis, appropriate quantities of the samples and reagents were mixed with standard solutions. The results of the analyses were calculated using optical reading at a specific time and temperature.
7. Histopathological examination of testicular tissues
For the histopathological examination, the testicles of the rats were removed and cleaned from the surrounding tissues. The testicles were fixed in a 10% formaldehyde solution for histological examination. The tissues were subjected to routine histopathological follow-up procedures after the detection process and blocked with paraffin. From the resulting paraffin blocks, 5-╬╝m sections were taken using a Leica RM2235 microtome (Leica Biosystems, Wetzlar, Germany). To analyze the complex histological structure of the testicles, the Crossmon triple staining technique was applied to the sections. The stained preparations were examined and photographed in detail with a Nikon Eclipse 50i research microscope (Nikon, Tokyo, Japan).
Results
1. Determination of TES, FSH, and FAS levels using ELISA
The levels of FSH, FSH, and FAS in the extracted testicular tissue supernatants are presented in Table 1 as mean┬▒standard error (SE). The average TES levels in the C, HC, and HCL groups were 172.34┬▒3.86, 121.75┬▒8.82, and 170.61┬▒6.59 (ng/mg tissue), respectively. The mean TES level in the testicular tissue was significantly lower in the HC group than in the C group, while the level was higher in the HCL group than in the HC group.
The FSH levels of the C, HC, and HCL groups were 129.41┬▒1.43, 115.47┬▒4.67, and 125.08┬▒1.28 IU/mg tissue, respectively. In the HC group, the mean FSH level was dramatically lower than in the C group; the level in the HCL group was higher than in the HC group and approached that of the C group. In the C, HC, and HCL groups, the FAS levels in the testicular tissues were 1.32┬▒0.06, 1.87┬▒0.08, and 1.51┬▒0.03 ng/mg tissue, respectively. The mean FAS level was significantly greater in the HC group than in the C and HCL groups.
2. Serum biochemical parameter levels in serum
The means and standard deviations of the serum LH, ABP, and total cholesterol (TC) levels of the C, HC, and HCL groups are presented in Table 2 as mean┬▒SE. The LH levels of the C, HC, and HCL groups were 12.15┬▒0.41, 7.77┬▒0.15, and 7.85┬▒0.43 (mIU/mL), respectively. The LH levels were lower in the HC and HCL groups than in the C group (p>0.05). The ABP levels of the C, HC, and HCL groups were 46.74┬▒0.76, 43.91┬▒1.72, and 47.12┬▒0.56 (nmol/L), respectively. Relative to the control, the ABP level was slightly higher in the HCL group and lower in the HC group (p>0.05). The TC levels of the C, HC, and HCL groups were 52.25┬▒1.75, 75.75┬▒1.98, and 60.95┬▒0.84 mg/dL, respectively. The TC levels were greater in the HCL and HC groups than in the C group.
3. Correlations
Correlations between groups with respect to TES, ABP, LH, TC, FSH, and FAS levels are presented in Table 3 as mean┬▒SE. The results included a significant negative correlation between TC and LH levels (r=ŌłÆ0.711, p<0.01), a negative correlation between ABP and TC levels (r=ŌłÆ0.282), a significant positive correlation between FSH and TES levels (r=0.535, p<0.01), and a significant negative correlation between FSH and TC levels (r=ŌłÆ0.656, p<0.01). A significant negative correlation (r=ŌłÆ0.723, p<0.01) was found between TES and TC levels. FAS level was significantly negatively correlated with LH, FSH, and TES levels (r=ŌłÆ0.548, p<0.01; r=ŌłÆ0.698, p<0.01; r=0.859, p<0.01), while it was significantly positively correlated with TC level (r=0.821, p<0.01).
4. Results of the histopathological examination
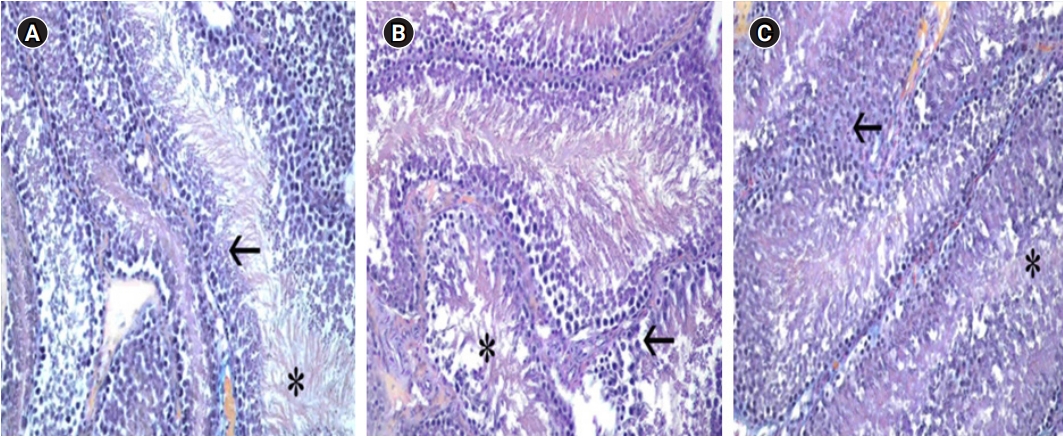
The testicular tissues were found to be surrounded by tunica albuginea, an externally irregular tight connective tissue. Seminiferous tubules of different lengths and diameters were observed in these structures, where the tunica albuginea divides the organ into septum by sending arms inside the organ, termed the septula testis. Spermatocytes and Sertoli cells at different developmental stages were observed in the seminiferous tubules. In the loose connective tissue between the tubules, small blood vessels and Leydig cells were detected. The tubule walls of the groups were evaluated and compared. In the testicles of the HC group, atrophy, vacuolization, and decreased integrity of the wall structure were observed in the cells in the tubule wall. A significant difference was found relative to the C group. However, in the samples from the HCL group, degenerate tubule wall cells were almost nonexistent, and the wall structure was preserved (Figure 1).
Discussion
The effects of probiotics on living beings and their health benefits have been emphasized in several studies. This includes their effect on hypercholesterolemia, particularly their role in suppressing advanced cholesterol synthesis and preventing the absorption of excess cholesterol [11]. Cholesterol is essential for the body because it plays a very important role in daily life. It is a crucial component of the cell membrane, as well as bile acid synthesis, fat and fat-soluble vitamin absorption, and synthesis of steroid hormones (sex hormones, mineralocorticoids, and glucocorticoids) [12]. A high-cholesterol diet is the primary cause of hyperlipidemia, atherosclerosis, and other lipid metabolism disorders that lead to male reproductive system defects [13]. In hypercholesterolemia, erythrocytes and endothelial cells, as well as serum, increase the amount of cholesterol present. The increase in the quantity of oxidized free radical products in these cells has been reported to cause elevated cholesterol [14]. Hypercholesterolemia is currently a widespread and common health issue. Several studies have shown that beneficial bacterial additives can lower serum cholesterol levels in fermented milk products or milk products containing lactic acid bacteria [10].The therapeutic value of fermented dairy products depends on the survival of these bacteria during development and storage [15]. In the present study, a rat model of hypercholesterolemia was developed, and L. acidophilus ATCC 4356ŌĆöthe lipid-lowering properties of which have been demonstrated in vitro and in vivoŌĆöwas administered intragastrically [10,16]. The results are consistent with previous reports suggesting that L. acidophilus has a cholesterol-lowering effect [17-19]. Walker and Gilliland [20] posited that L. acidophilus secretes bile salt hydrolase to deconjugate bile salts, although no connection was found between the reduction of in vitro cholesterol and the degree of bile salt deconjugation [21]. These conflicting findings raise the possibility of other mechanisms that may be associated with the assimilation of cholesterol by probiotic bacteria during their development [22]. In the present study, the TC level was elevated in the HC and HCL groups (p<0.05) relative to the control. Our analysis indicates that L. acidophilus plays a major role in cholesterol synthesis. The administration of this probiotic may result in increased catabolism of cholesterol in the liver, contributing to a hypocholesterolemic impact. Several clinical and epidemiological studies have shown an inverse association between serum TES levels and TC [23]. In addition, animal experiments in TES-deficient male mice have also shown dramatically elevated serum cholesterol levels [24,25]. Circulating TES levels are determined by the steroidogenic potential of Leydig cells and the total number of Leydig cells per testicle. Stress-induced increases in serum glucocorticoid concentration have been reported to decrease the rate of TES release by inhibiting the activity of TES biosynthetic enzymes [26]. In another investigation of the impact of probiotics, larger testes and higher serum TES levels were observed in male mice that regularly ingested lactic acid bacteria than in age-matched control mice. Lactobacillus reuteri was administered as a probiotic in that study, and a thorough analysis with microscopy-assisted histomorphometry showed an increase in the extent of spermatogenesis and the number of Leydig cells per testis. Furthermore, gonadal aging symptoms were claimed to decrease [27]. Mega Acidophilus (a mixture of L. acidophilus, Bifidobacterium bifidum, and Lactobacillus helveticus) was administered to New Zealand rabbits for 1 month to determine how the probiotic would affect certain parameters. In that study, probiotic supplements were shown to improve antioxidant activity and substantially increase the level of TES [28]. In our study, the TES level in the group given 2% cholesterol for 8 weeks decreased in the testicular tissue, and the level of TES increased in the group given the L. acidophilus probiotic. Therefore, TES deficiency in the liver in HC does not affect de novo cholesterol synthesis. A number of human studies have shown that hypercholesterolemia and low semen quality are associated with male infertility [29-31]. Research on animals fed a high-cholesterol diet has uncovered more information about this link. In animals with diet-induced hypercholesterolemia, adverse effects of hypercholesterolemia on testicular function, including spermatogenesis, were noted [5,32-35]. Increased oxidative stress is of considerable research interest among the many pathways suggested for hypercholesterolemia-induced testicular injury [16]. When mice were fed a high-fat diet, decreased TES, decreased semen quality, seminiferous tubule atrophy, and degeneration were reported. The researchers stated that the addition of selenium-enriched probiotics to the high-fat diet decreased damage to testicular tissue and raised serum TES levels [36]. In another study, researchers examined the reproductive system structure of male rats fed a high-fat diet for 2 months. They found that sperm quality and count decreased, Leydig cell disturbances increased, spermatocyte and spermatid structures decreased, and significant narrowing of the seminiferous tubule occurred [37]. In our sample, relative to the control group, the testicles of the group given cholesterol feed (HC) showed atrophy, vacuolization, and damaged wall structure integrity in the tubule wall cells. However, in the HCL group, degenerate tubule wall cells were almost nonexistent, and the wall structure retained its integrity. According to recent research, high cholesterol levels caused by high-fat diets contribute to urological disorders (problems with penile erection, irregular spermatogenesis, benign enlargement of the prostate, cancer, etc.). They also disrupt the epithelial structure of the tissues and cause functional disability [11]. The secretion of FSH and TES is required for the successful completion of spermatogenesis. TES, the male sex hormone, is secreted by Leydig cells under LH stimulation and plays important roles in the differentiation of peripheral tissues and the promotion of spermatogenesis [38,39]. ABP binds to TES and estrogens and assists their transport into the seminiferous tubule, allowing for their use for spermatozoon maturation when required [40]. Testicular ABP synthesis has been reported to be increased by FSH and TES in rats [41]. In another study, researchers investigated the impact of Lactobacillus rhamnosus PB01 on sperm kinematic parameters and found that TES, LH, and FSH levels, as well as sperm motility rates, were all significantly elevated. L. rhamnosus has also been found to act as a positive regulatory agent on weight loss and reproductive hormones [42]. In our study, serum ABP and LH levels were lower in the HC than in the control group, and the amount of FSH in the testicular tissue was also lower. ABP and LH levels were greater in the HCL group than in the HC group (p>0.05). In parallel to our results, when a 2% cholesterol diet was given to male Wistar rats for 21 days, non-significant changes were seen in TES, LH, and prolactin levels, while the FSH level was significantly decreased [43]. In another study, a high-cholesterol diet induced a significant increase in the TC level and significant decreases in the FSH, LH, and TES levels in the serum of male rats [44]. Hypercholesterolemia has also been suggested to cause reproductive and testicular damage through excessive free radical generation and increased oxidative stress, which is cytotoxic to spermatozoa [13,34,45]. In hypercholesterolemia, the administration of antioxidants and lipid-lowering agents has been shown to protect the testis and reproductive functions [13,34,46]. In the present study, the FAS level was significantly greater (p<0.05) in the HC group than in the control group, while the FAS levels did not differ significantly between the HCL group and the control group (p>0.05). The FAS level was significantly negatively correlated with LH, FSH, and TES levels, while it was significantly positively correlated with TC level.
This study revealed that probiotic treatment is beneficial for reducing the cell degeneration in testicular tissue caused by high cholesterol. Probiotic treatment increased LH, FSH, TSH, and ABP levels, while partially reversing the increase in factor-related apoptosis associated with a high-cholesterol diet. Probiotics have an important role as a supplementary treatment for degeneration and as an adjunct in the treatment of cell structure disorders, although they are not therapeutic in isolation.